����������:
1) ������������� ����������������� ������ ��������� �������� �� ������ ������� ������������ � ��������� �� ����� � ������������ ���� (���������� �����������, ������� - �.�. ���������);
2) ����� ����� �� ����� ��������������� �� ������������� ��������, ���������� � ������;
3) ������ ������ �������� ��������� � �� ����������� �������� ������ � ��������� ������� � �������� ������������ �� ���� www.ecosystema.ru
������ ������ ���� ������������ � ������� �������������� ������ ������������� �����
"������� ����"
�������� ������������ ���������
������ � ������� �������������
����������� ��� ���� � �������
�������������� �.�.,
������������ �.�.
1997
����� ���� ������� � ������� ��������
�������� ��������������, �� ������, ��� ��������
� ������ � ��� ���������������� ���������� ��
���������� �������������� �� ������� ����������
� ������� ���� ���. � ���������, ���� ����� �����
���� ��������� ���������� � �����������
������������ ��������� � ����������� �������
��������, � �� � ����� ���, ������� ���������� ��
������� �����. ����� ����, ��� ������ �����������
�� ������� �������������� �� ������ �� �����
����� ����������, ��� ��� ������� � ������������
������������� ���������� �� ���� ����� ��� ��
����������� ������� ������������� � ���������,
����������� � �.�. � ����� � ���, ��� ��������
�������� � ���������� ���������. ����� �������,
�� �� �����, � ����� ������� �� ��� ���� ���������
����� �������� ������������ � � ����� �
����������. ������ �����, ������ �� ���������
����� �������������� � ���������� ���������
����� ��� �������� ����������� ����� �����.
������, �� ������ �� ��� ��� ����������, ������
���������� �� ������� ��������������
������������ ����� ������� �������. ��� ���� ���
������� �������� �����������, �������� ������� �
��������, � ����� ������� ��� �������� ����
��������� �������������, ���������� ����������
��������.
� 1985 ���� �� ���������� ����������� ������� ������� ����� ���
���� ��������� ������������ ��������������
������� ����� 5�10 �� � �� �. ���� �� �. ������� � ��
���������� ������ ������-��������� (��������
���) �� ���������� ������ � �. �������. � ������
��������� ������������ ���������� ���
������������ ���� �������� ������������� �
������������� �������� �����������,
������������� �.�. ����������� (1981). �������,
�������� �� ������������� �����, �������� ��
��������� ������������ ��������� � ������� �
������������� ����� ��� ���������� ��������
������ ��������� ����� �����, ��� ������ ����
�������� �����. ����� ������� ���� ��������
���������� ������ ������������ ���� �� �����
��������������� ���� ��������������. �����
������ ���� ����� ���� �������� �� ����� �
�������� 1: 25000. ��� ����� ���� ������������
��������������� ����� � ����� ������������ 5 �.
������������ ����������� ���������� ������� �
�������� �������� �������� ������� ������� �
���������� �� ����������, ������ �� �����
������� ������������ ����������. �����
������������� �������������� ����������
���������� �� ������� � ����� �������������
�������� ������ ������ �� �����. ����������
������� ��������� �����-����� ������������
������������� ������� ������. �������������
������ � ���������� �����, �������, ����� � ���
����������� ����� �� �����������. ������
��������� � ����� ������ ������� ��������
������������ ���������, ������������� ��
���������� ������������ �. ����, �
���������������� �� ������������� ��
��������������. ������������ ��� ������� ���
�������� � ��������. ��-������, ��� �������� �
������������ ������������ ��������� ���������
������������ ����������, ����������� ��-�� ����,
��� ������ ���� �������� (���, �����, �������,
�������� � �.�.) ������ � ������ ���������� ������
������������ �����. ��-������, ����������
������������ ���������, ������� ���������
������ ���������������� � ����� ������
������������� �����, �������� �������
��������-��������� (�� Sphagnum girgensohnii). ���������
�������� � ��� ������� ������������� ���������
������������ ���������� �� �������. ��� ���
������ ��� �� �����, ��� � ��������
�������������� ����������� ���� � �������
������ ���� �� ����������� �����������. ���
����� �� ������ ��������� ��������
��������������, ��������� 13 ��� �����, � �������
������������ ���������. ������������ �� �������
����� ���������, ��� � 1984 �., ������
�������������� ��������������. � ���������
������ �� ���������������� ��� ������������
���������, ���������� ���������� ������� ��
��������� �����������.
��������� � ������
� ���� ������������ 1985 �. ���� ������� ����� 160
�������� ��������������. ��� ����� ������
�������� � �������, �������� � ������ ����������.
����� 1997 �. � ����� 13 ��� � �� ��������� �����
���� ��������, � ������� ������������ �� ����
���� ���������.
��� ����, ����� ���������������� �������
�������� ��������������, ����������� ��������
���� � � ������ ����� �������� �����������
��������� ������. �������� �� ���������� ������,
�� �������� ��������� ����� ���� ��������������,
������������ � ������������ �����
�������������. � ��������� ������ �� �����������
������ ������������ ���������� ������ � �����
�������������. ��� ����� ������������ ���������
28 ��������.
�������� �������������� ��������� �� ��������
�������� 10 �. ��� ���� � ������ ����� ���������
����������� (����������� ��������) ������� ����
� ��������� �� ������ ��� ����� ����������. ���
�����, �������� ������� ���� ����� 10%, ���������
������ ������� ��������: 1-9% (������) ��� ����� 1%
(���������). ��� ���������� �����, �������� �
�������� ��������� ����� ������ �, ��
�����������, �������.
��� �������� � ���������� ����������
������������ ����� �� ������������ ��� ����
����� �������� ���� �������������� � ���������
������������ ��������� � �������
��������������. �������� �����������,
������������ �������������� ������� ��������
�������������� � �������� ���������� �
���������� ��������, ������ �����. ��������, ���
� ���� ��������� ���������� ��� ������������
�������������� ������������ � ������ �������
������� ��������� ����� � ��������� � �������
����� ��������� �. �������������, ���� ������
������������ ���������� �������� �������������
�� �������� ������������� ����� � ������
����������, �� ���, ������ �����, ������������
����� ������ ������ � ���� �� ��������������
����. ��������� ���� ��������, �� ��������
���������, �� ������������� ����� ���� � ������
������������ ����������, ����������������,
�������������� ����� ������ ����� � ��������,
������ ��� � ����������� ������� �� ���������.
����� ��������� � ������ �� ��� ����� � �����
��������� ������ ���������� � �������������� �
������ ��� ������ �����. ��� �� ��������� � ���
������, ����� � ���� �������������� ������
���������� �������� � ������ �����������
���������������.
����������
�������������� ������ �������������
�� ���������� ����������
� ��� ������ �������� ������� � ������������
������������ ������������ �������� ����.
������� ���� ���� ����� ����������� �������,
������� � ����������� ����� � (���) �������
��������. � �������� �������������
������������� �� ������� 40-60 �� ������������
������������ ����������������� ��������; �
��������� �������������� �� ��������� �� ���
(����� ������������ ��� ���������� �� �������
�������). ��� ���� ������������� �������� ����
���������� ���������� ���������� (������, 1980).
������ � ����������� �������� � ������ ���������
(���� ��������� �) ���� �������� ��������.
� ������ �������������� �� ����������
���������� ����������� � ��������
������������������ ����� ������ �������
�������������. �������� ��������� ������
���������� ����� 5 ��. �� ������� ������� ���,
��������, ������� ���������� (�� 6 ��), ������
������ ��� ������ ���� ��������� ������������.
�������� ������� � ���������
����������, ��������� �����, ����� �������
������-���������. � ��� ��������� � ���
������������� ������ ������������ � ��������
(Betula pendula, B. pubescens), �������� ����� (Populus tremula), �
�������� � �������� �������� ��� (Picea abies), �����
(Abies sibirica), ������ (Sorbus aucuparia), ������� ������ (Frangula
alnus). � �������-�������������� ����� ����������
������� (Oxalis acetosella), �� ���������� � �������
(Vaccinium myrtillus). ������������ ���� ��������
���������� �� ���� ���������� ����������� �
���������� ������������� (Stellaria holostea), ������
���������� (Majanthemum bifolium), ��������� ����������
(Trientalis europaea), �������� ����������� (Asarum europaeum),
���������� ������ (Gymnocarpium dryopteris), ��������� (Rubus
saxatilis) � �.�. ������� ���� ������ �����, ���
�������� � ����� 10 %.
������� ������ �������� � ������,
���������� ����� 13 ���, �� �������� �����
������������ ��������� �������������� (���.1).
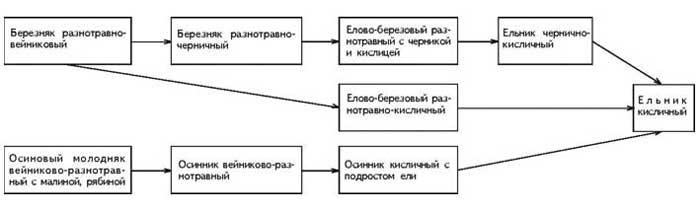
���. 1. �������� �������������� ������
������������� �� ���������� ����������
���, �� ����� �����������-�����������
�������� ��������� (�� Betula pendula, � ���������
�������� B. pubescens) � �������� ������ (Calamagrostis
arundinacaea) ����������� �������� ������������ �
�������� (����������� ����� ������������
������� ���������� ����� ������� � �������
������ ��������� ���������� � ������������
����������� ������: ������������ ������������
(Solidago virgaurea), ���������� ���������� (Dryopteris spinulosa) �
�.�.). ���������� ������� �������� ���������� ��
���� ����� ����� � ����� � ����������� ���������.
������������ ���� � ���� �������� ������ �����
�������� ������� ���� � ��������. �����, �� ����
����, ��� ��� ���������, ����������� ���������
�����-��������� �����������-��������-���������
���. � ���������� ��� ��� ������� ����������
��������-�����������, �������� �����
�������������� ������ ��������-���������.
����� �����������-����������� ��������� ��
������-��������� � ������������ ����� ���������
� ����� ��������-��������� ������. ����� ������
������� ���� � ��������, �������� �������������
��� ��������������: �� ��������� �������������
��������� ������� � ���������� ������ ��������
������������� ������������ ���������.
� ��� �� ����� �������������� ����� �������
����� ������������� �� ���������, � ��������
���������. ���, �� ���������, ��� �� �����
��������� ��������-������������� c ������
���������, ���������� 13 ��� ����� � ������������
����� ���-��� ����� ��������, �������������
������� ���������-������������ ������� ��
������������ �������� ������ � ������ � �������
��������-�������������� �����. �����
������������, ��� �� ��� ����� �� ��������
������������ �����������-���������� �������.
����� �������� ������ ���� �� �������-���������
� ������������ ��� ������� ������������.
��������� �����, ����� ���� ������� ��������
��������������� ��������-�����������, �����
��������� ������.
����� �������, � ��������� �����������
�������������� �� ���������� � �����������
������ ����� ��������� ��������� �����
��������������. ��������� � ����������� ������
������������ ��� ����� ��������� ������. �
�������� ������� ������� ������������ ������
������, ������� �� ���� ����� ���� ����������
�������� � �������� ��� ���������������
��������. �� ���� ������� ������������ ���� �
������������ ������� ������ ���������
�����������. � ������� � �������� ����������
�������������� ������ ����������� ������ ������
���������� ������������� � ��� ����������� (Milium
effusum); �� �������� ����������, ����� ����,
������������ �������� (�� 10%) ����� ����������
��������� �������� (Melica nutans) � ���������. �
��������� ����� ���������� � ���� ������ ���
������, �� ��������� � ���������� ����� ���� ���
�������� �����. � ������ �������� ������
����������� �����������, �������� ����� �����
��� ����� �������� ������. �� ��������
���������� ������ �������: ����� ������,
����������� ����-���. ������� ������
�������-��������������� ����� � ��������
��������� �� ���� �������. ���, �� ������� ���
������� ������� � �������� ������ ����� �����
������� � ������ ���� ������ �����, � �������, �
����� ����� ����� � ��� � ������ ������. �������
���� �� ���� ������� ����������� ����� �
��������� � �������� ����� 10% ������ � ��������.
� ����� ������� ��� ���������� ����,
�������������� ����� ��������� ��� �������
���������� ����� ��������, ����� ���� ���������
�����. ����� ������ ������������ ������ � ������.
��� ��� ���� ������ ������������ � ���������
����������� ��� ������� ���������, �� �����
���������� ����� ����������� ���� ������� �
��������. � �������-�������������� �����
������������� ����������� �������� ����,
���������� ������������� � ������� �������, �� �
������� � �������� ����������� � ������������
�����������. ���������� ���, ������ � ��������, �
���� �������� ������ �����. � �������������
������� ����� ��� ����� ����������� �������
�����.
�������������� ������ ������������� ��
������
�������� ����� ������ ������������ ��
���������� ���. � ����������� �� ������
��������� � ������� ������������ ������������
������ �� ��� ����������� ������ ������������
����������. ����� � �������� �� �������� ������
��� ���� �������� ����������� � ������ �.
��������� (1996, 1997). ��� ��������� �������������
������������� �� ������ ��� ������� �����
������������ �������� �� ���������-������������
������� � ����� �� ������� ������������.
���������� ������ ���� �� ������������� ����
������� ���������� ���� � �������
�������������� ����. ��� �������� ������� ������
� ����, ��� �������� �����, ������, ��������
������. ����� ������������� �� �������������
������� ������ ���, ������������� �� ��������� �
��������� �. ��������� ���� �� ������� �, ������,
������������� � �������� ����� ��������
��������� ������������ ���������. ��� ���� �����
���������� �������� ����� ������ �������
��������������. ������ ��������� � � ���������
������� ������ ���������� �������� ��������.
�������� ���������� ����� ���� � �������
��������-������������. � ��������� ����� ���
����� ����� ����� ����� � ������ ����� �����.
������� � �������� ������ �����, � ���
����������� ���, �������� ������, �������
�������, ������, ������������ (Juniperus communis). �
�������-�������������� ����� ������������
������� (������ ����� 70%), ��������� �����������
�������� (Vaccinium vitis-idaed), ������ ����������, ������
�������� (Linnaea borealis), ��������� ������� (Melampyrum
pratense). ������� ���� � ��������� ����� 30% ������� �
�������� �� Pleurozium schreberi, � �������� �������������
Kicranum polysetum � Hylocomium splendens.
� ������ ������������� ��������������
��������� ���� ���� ������������ ��������� (���.
2).
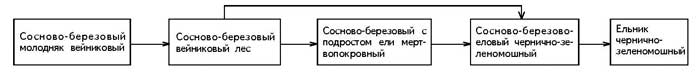
���. 2. �������� �������������� ������
������������� �� �����
����� ����������� ��������� (�� �������)
��������� ��������-�������� ��� ���������
�������� (� ������������� � ����� ������� �. penduld).
� ��� ����������� ������� ���������� ������
������. �� ���� ����� ����������� ��������
���������� �������� ��� �������-��������� ���.
����������� ������ ������ ���� ����� ����� ��
������ ���������. � ��� ������������ ������
������, � ��������� ����������� �����
����������� ��������, ������� ����� (Antennaria dioica),
��������� ������� � ������ ���� �����������.
���������� ������� ���������� ������� ���. ��
���� ����, ��� ��� ������� � ������ �����������
������ �������, ����������� ������ ��������:
������ ������� �������, ������� ���� ����������
������. �������, ���, ��� ��� �������� �����, ��
����� ����������� ���� ������� ���������� ��� �
������ ������ ��������� � �������������
�������� �������. � ������ ��� � �����������
����� �� ������������ � �����������-���������
��������� ��� �, �����, � �����������-���������
������.
� ��������� ���� ��� ������� ����������
��������� ������� ����� ����� � ���� �����
��������������. ����� �������������
������������� ����� ���������� ��� ������
������ ���� � ���������� �����. �
������-��-������������ ���� ��� ��������� ��
������������� ����� � ���������� ��������� 4 ��,
��=4 ��. � ��������� ���� �� ������� �������������
����� � ���������� ��������� ����� 6 ��, ��=4 ��. �
������� ������� ������������� ����� �
���������� 7 ��, �1�2=8 ��. �� �����, ��� �����
������������� ����� ��������� �� �������������
�����. ����� ���� ����������� �������� ���������
� ������������ ��������� �1�2.
�������������� ������������ �������������
������������� �� ���������� ����������
� ���� ��� ����� ������������ �������������
������������� � �������� ����� �� ������������
�������� ����, ��� � � ������ �������������� ��
������� � ���������. ������� �������� ���� �
�������� ��� ����������, ������ � �����������
���������. �� ������� 40 � 70 �� �������� �������
������� ��� ������� ��������, ��������
����������. � ��������� � � ������, � ������ � �
��������� � ����� ������� �������.
������������ ������������ ��������������
����� ���� ������������� ������ ��������� �����
Sphagnum girgensohnii, ���������� �� ���� ����������� ����
10 � 30 % ����������� �����. �������� ������� �
��������� ����� ������������� ��������� ����
�������� ������ ��������-���������. � ���
��������� � ��������� ����������� �����������
����� � ������ ����� �����. ������� � ��������
������� �����, � ��� ������������ ���, �
��������� ���������� (������� ������, ��� �
������������� ��������������) � ������,
�������� ������� � �����. � �������� �������
���������� �������� � ������� (�� 80% ��������).
��� ��������� � ���� �������� ������� �����,
������������ �� ����� �������� �� ��������
������������� �������������. � ������� � �����
��������� ����������� ������������� ������� �
���� ���������� ����������� � ������,
���������� ������������� � �.�.. � ������� �����,
����� ��������, � ��������� ����������� (�� 10% �
�����) ����������� Pleurozium schreberi, Kicranum scoparium � Hylocomnium
splendens. ������ ���������, �� ������� ������ ���,
�������, ��� � ����������� ����������� �������
��� �������� ��������������, �������������� ��
����, ���������� ������� ��������, �.�. ������ ��
�������� ���������.
����� ������� �� ����� ������� ���� ���������
���������-��������� (� �������� ������) �������
��� ��������, � ������� ��� ���� �����
������������ �������� � ������ ����������� (���.
3).
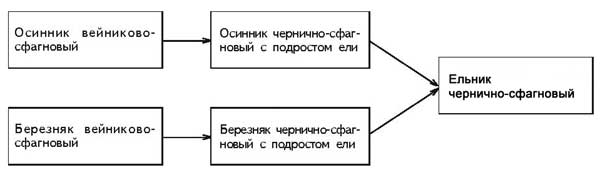
���. 3. �������� �������������� �������
������������� �� ��������
����� ��������, ��� ������ ������
������������� ��������� ���������, � �� �����
��� ����� � ������ ������ ����������, ��
�������������� ����������. � ������ ��������� �
������������� ������� ����� ������ ���������
��������, � � ����� ������ ����������� ���
��������� ������ ��������-���������. ���� � ����
������� �� �����-�� �������� ���������� ����, ��
� ���, ��� ��, ��� � ��������� �������������
�������������, ������������ ������ � � ���������
����������� ������. ������������ �����������
������������� �������� ��������. ������������,
���� ������� ���������� �������� �������� � ��
��������. ������ ���� ��������� �� ��������� ���
������������.
� ����������� ������������� �����������
������ ������������� ��������� �������� ������
8-10 ��. ��������� ��������� � ����� � ���� ����
����� �� ����� ������ �� ��������������.
��������� � ����� �� ��������� ��������
����������� ���������� �� ����������� �
���������������� �������� ��������, ��� ��� ���.
�� ��������� ������ ��������� ����� ������� �
������������; ��������� ������� ����� ����
��������� ���������, ������ ��������� ���������
� ��� ����� ���������� �� ����. �����
������������, ��� ��������� ����� �����
���������: ��� �����������, ������� �����������
������ ����� � ��������� �������
����������������, ��������� ��������� ���������
��������� � ��������� ���������. ��� ����
��������� ����� ����� ����������� ���
�������������, �� �� ��������. ��� ������������
������ ��������� ������ ��������� ����� � �
���������������, ��� ������� ���. ������, ������
�������� ����� �������� ������� ���������
������������.
����� �������, ��� ������� ������� ��������
�������������� � ���� ������������ ����������,
���������� �������� ����� ����������
����������� ��� ���� � �������. ���� ���� � ����
���������� ������������ �������� �� �������
�����-���������� �������� ������
��������������. � �� �� �����, ��� �����������
���������� �� ��������� ���� �, � �����������,
���������. ��� ����������� ����� ��������������
� 1984 �. ������ �� ������������� ����� ��
��������� ��� �������� ������������ ����������
������� ������ ��������� ���������� � ��������;
������ � ��� �������� ������� ������
����������� ���������� � ��������. ����������
���������� ������������ �� ���� ������
��������� ������ ����� ���������. ������ �� ���
� �� ����������� ������� ������ � ��
����������� ��� �������� ���������� �������� �
��������� ����������, ��������������
����������� ������������ �������������. � ����
������������ ����������� ������. ��-��������,
��� �������� ��������-��������� �� �������
���������� ������ �������� ����������
���������� �������-������ ����������.
����������
- ����������� �.�. �������������� ��������
����������. �. �����. 1981. 240 �.
- ������ �.�. �������� ������ ������. �. �����. 1981.
190 �.
***
������ ������ ���� ������������ � ������� �������������� ������ ������������� �����
"������� ����"
������ ����������������� ������, ��������� � ������������� ������ ����, ����� ����������
� ������� ����������������� ������. ������ ������� ����������������� ������ (����������������� �������), ����������� �� ����� �����, ����� ����������
���.
 ������� ������
"�������� ����"
��������� "Parus"
������ ���������
������� ������
"�������� ����"
��������� "Parus"
������ ���������
|
���� ��������� ������������ ��������� �� �������� � ��������� ������:
� ����� ��������-�������� �� �������������� ����� (�� ������������� ������������)
����� ���������� ��������� ������������ ���������
�� �������� � ��������� ������:
 ��������� ������� ������������-��������:
��������� ������� ������������-��������:
 ������� �����,
������� �����,
���������� �����,
���������� �����, ������� �����,
������� �����,
���������� �����,
���������� �����,
������������ �������� (��� PC-Windows) ������������:
�����,
������� �����,
������� �����,
����� � ������ �����,
����������-������������ �������� ��� ���������� � ��������� �� ��������:
�����,
������� �����,
������� �����,
�����
(�� ����� ������� �� ��������� ���������� �� ������� �� ��������� ��������),
����������-������������ �������� ��� iPhone � iPad:
������� �����,
�����
(�� ����� ),
��������� ������� ������������:
������������� ��������,
�������� �����,
�������� ������� ���,
������� "������ ����������������� ������������",
������� �������������� ��������������� �������:
�����-1,
�����-2,
���������,
���������,
���,
����������,
����� �����,
����� �����,
����� ��������,
������� �����,
������� �����,
���������� �����,
���������� �����,
������������ ����� "������������ ������� ������"
������� ��������.
|
|

