����������:
1) ������������� ����������������� ������ ��������� �������� �� ������ ������� ������������ � ��������� �� ����� � ������������ ���� (���������� �����������, ������� - �.�. ���������);
2) ����� ����� �� ����� ��������������� �� ������������� ��������, ���������� � ������;
3) ������ ������ �������� ��������� � �� ����������� �������� ������ � ��������� ������� � �������� ������������ �� ���� www.ecosystema.ru
������ ������ ���� ������������ � ������� �������������� ������ ������������� �����
"������� ����"
������������� ������ �����������
��������� ��������� � ������
����������� ���� �������������� �������
��������-����������
�.�. ������, �.�. ��������, �.�. ��������
1999
��������
�������� �������� ��������� ��������������
� ���� ��������� �������������� ���������
������������ ����� �����. � ���� ������������
������ ������� �������� ��������� � ��������
���������� ���������������� (�������, 1979;
����������, 1979; ������, 1986). ������, �� ��� ������,
������� ��������� ��������� � ����
�������������� �������������� ������ ������
������������� ��� ������������ ����. � ����
���������� ������ (������, �����������, 1997) ����
���� �������� �������������� ����� �� ������ �
����� ������������� ��������, �� � �
������������ ����� ����. ������������ �����
����� � ��������� ������ ������.
��������� � ��������
������� ������������ ����������� � 25 ���� ��
26 ������� 1998 ����, �� ���������� �����������
������ ������-������� ������� ���� ���,
������������� � ������������ ������ �����������
�������, � 30 �� �� ������ ���������.
��� ������������ ���� ������� 5 �������������,
�������������� ����� ��� �������������� �������
��������-���������� �� ������� �����������
������ (��������������, ������������, 1997) (���. 1).
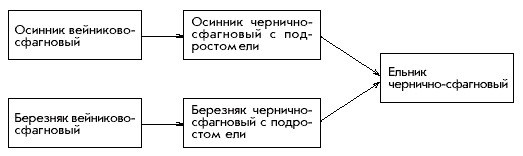
���.1. ��� �������������� �������
��������-���������� (��: ��������������,
������������, 1997)
������� ���������-���������. �
��������� (����������� 0,6) ����������� ����� (60%),
������������ ��� (20%), ����� � ������ (�� 10%). �
�������� � �������� (����� ����������� 0,3)
��������� ������ �������� ��� (50%) � ������ (30%). �
�������-�������������� ����� (�����������
�������� 60%) ����������� ������ ������ � �������
(�� 15%). ������� ���� (����������� �������� 90%):
������� � �������� �� Sphagnum girgensohnii (60%).
������� ��������-��������� � ��������� ���.
� ��������� (����������� 0,6) � ����� (60%), ��� �
������ (�� 20%). � �������� � �������� (�����
����������� 0,3) ����������� ��� (70%). �
�������-�������������� ����� (�����������
�������� 50%) ����������� ������� (35%),
������������ �������� (10%) � �������� ����������
(<10%). ������� ���� (����������� �������� 60%): Sphagnum
girgensohnii (50%), Sphagnum riparium (<10%), Pleurozium schreberi (<10%).
�������� ���������-���������. �
��������� (����������� 0,7) ����������� ������ (80%),
����� ������������ ��� (20%). � �������� � ��������
(����� ����������� 0,5) � ������ (20%) � �������
������ (10%). � �������-�������������� �����
(����������� �������� 65%) �������� ���� � ������
������ (20%), ���������, ���������� ������������ �
�������� ���������� (�� 10%). � ������� �����
(����������� �������� 40%) ����������� Sphagnum girgensohnii
(20%).
�������� ��������-��������� �
��������� ���. � ��������� (����������� 0,7)
����������� ������ (70%), ����� ������������ ���
(20%), ����� � ����� (<10%). � �������� � ��������
(����� ����������� 0,3) ����������� ��� (80%). �
�������-�������������� ����� (�����������
�������� 55%) ����������� ������� (35%),
������������ ����� �������� ������ ������ (10%). �
������� ����� (����������� �������� 10%) � Sphagnum
girgensohnii (60%).
������ ��������-���������. � ���������
(����������� 0,6) ����������� ��� (80%), ������������
����� � ������ (�� 10%). � �������� � �������� (�����
����������� 0,2) ���������� ��� (80%). �
�������-�������������� ����� (�����������
�������� 60%) ����� ������� (40%) ������������
�������� � ��������� ������� (<10%). ������� ����
(����������� �������� 60%) ����������: Pleurozium schreberi
(60%), Sphagnum girgensohnii (35%), Hylocomium splendens � Sphagnum russowii (<10%).
���� ���� ��������� ���� ���������, �� ������ �
������ ����������� ���� ����. �� ����� ��������
�� �������� �������� � ��������� ��� ��������
���������-���������� 80 �, ��������
��������-���������� � 200 �, ����������
����������� � ���������� 100 � 70 � ��������������, � ��� ������� � 300 �. ������
10 � �������� ���������� ��������. �� ����������
������� �������� ���� ������� �����
��������������� ��������.
����� ������� ������� (�� ����� 2-3 �), �� �����
����������� ��������� ����� �������� 0,0625 � 2.
��� ����� ������������� ������������� ���������
������� �� �������� 25 � ������� 15 ��. �����
����������� �� �����, � ��������� �������������
� �������������� ����� � ����������� �
������������ �������� �� ����������� ��������
(�������, 1975; ����������, 1975). �����������
��������� ����������� �.�. ��������.
��� �������������� ��������� ���������
��������� ������������� �������� ���
����������� ��� �������������.
�������� ����������� ������� �
�������������� �� �������: K = C / (A - B - C), ��� � �
����� �����, ����� ��� ���� ������������
�����������; � � ����� ����� � ������
�����������; � � �� ��, �� ������. ���������� � ��
������� ������������ ���������� �������
�������� �������� ������� ��������� � ����� �
������ ������ ���� ������ ��������.
���� �� ����� ����������� � �����������
�������� ��������� ������, ������������
����� ����� ����������� �������� ��������
������ ������� �� ����� (������,1964)
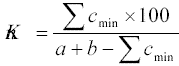 , ,
��� � � ������� (�� ����) ���������� ������
������� ����, � � ��������� ������ ���� ����� �
����� �����������, � � �� ��, � ������. ������
���������� �������� ����������, ��� �������
������������� � ����� �������� �������
��������������� �������� (������,1975).
����������� �.�. ����������: K = (Kn x Kf) / 100,
��� Kn � ����������� �������� ���������
������, Kf � ����������� ���������������
��������, �������������� �� ������� �������.
������ �������� ������������� ����������
�������������� ������ ����, ���������� � ����� �
��������� ���������, � ����� ��������� �������
����������� ��� �.�. ���������, ��������� ������
� ����������� ������.
���������� � ����������
� ����� ��������� ���� ���� ��������� 150
����������� ��������������, ����������� � 19
����������, 33 ����� � 75 �����.
� �������� ���������-��������� ���� ���������
16 ������ 13 �����. ���������� �������� ���������
��������� (Rhagio sp.) � 6 ������/�2. �� ������
����� � ������ ��������� (Athous subfuscus Muell.) � 4
�����/�2 (����. 1).
������� 1. ������� ������ ���������
��������� (������/�2)
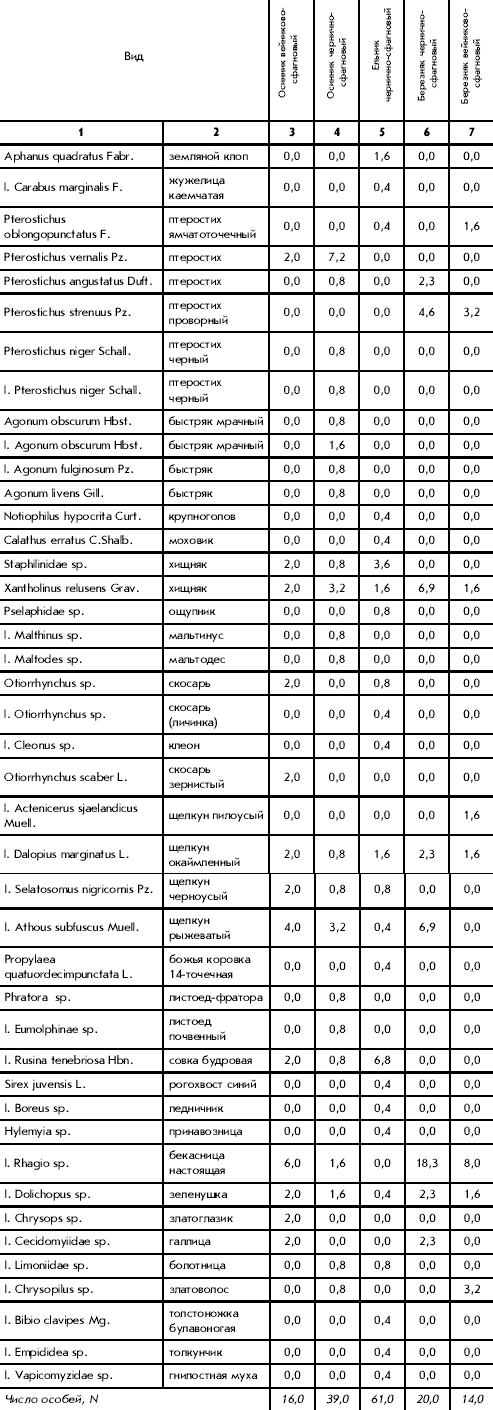
� �������� ��������-��������� ���� ����������
39 �����������, ����������� � 22 �����. �� ���
���������� Pterostichus vernalis Pz. (7,2 �����/�2).
� ��������� ���������-��������� ���� ���������
14 ������ 8 �����. � ������ ������������� � �����
���������� ������� ��������� ��������� (Rhagio sp.)
� 8 ������/�2.
� ��������� ��������-��������� � �����
�������� 20 ����������� 8 �����. ��� �������������
��������� ��������� (Rhagio sp.) � 18,3 ������/�2
� �������� ��������� ���� �������������� �����
� ������� ���������� (Athous subfuscus Muell.) � �������
(Xantholinus relusens Grav.) � �� 6,9 �����/�2.
� ������� ��������-��������� ������� 61 ����� 24
�����. ���������� ���, ����������� � ����
��������� � ����� �������� (Rusina tenebriosa Hbn.) � 6,8
������/�2. ������������: �������� ����
(Aphanus quadratus Fabr.) � 1,6 ������/�2 � ���,
����������� ��� ������� �������������, � �������
(Xantholinus relusens Grav. � Staphilinidae sp.) � �� 1,6 � 3,6 ������/�2
��������������.
� ���� ��������� ����� ���� � ��� Pterostichus
vernalis Pz., Xantholinus relusens Grav., ��������� ��������� (Rhagio
sp.), ��������� (Dolichopus sp.) � ������; ������
����������� (Dalopius marginatus L.), ������ ���������
(Selatosomus nigricornis Pz.), ������ ��������� (Athous subfuscus
Muell.), ����� �������� (Rusina tenebriosa Hbn.) �
����������. � �������� ���������-���������
������� ������� ��������� (Otiorrhynchus scaber L.),
��������� �� �����. ������� ����� ���� ����
��������� ������ � ������ ��������. �����
��������������, ��������, �����������
������������� �����, ���� � �����������
��������� ���������� ������� �������������.
� ���� ���������� ������ �������� ���������
��������� (Pterostichus strenuus Pz.), Xantholinus relusens Grav., ������
�����������, ��������� ���������. ����������
(Chrysopilus sp.), ������ �������� (Actenicerus sjaelandicus Muell.),
��������� �������������� (Pterostichus oblongopunctatus)
���� ������� � ��������� ����������, � Pterostichus
angustatus Duft. � ������ ��������� (Athous subfuscus Muell.) � �
��������� ���������.
� ������� ��������-��������� �������� ����
�������������� ������ ���������: �������,
������� (�����������, ���������, ���������),
����� �������� � �� ���������, � ���������
�� ����������. ������ ���������� � ����� ����: ��������
���� (Aphanus quadratus Fabr.), �������� ��������� (Carabus
marginalis F.), ������� (Calathus erratus C.Shalb.), ��������
(Pselaphidae), ����� ������� 14-�������� (Propylaea
quatuordecimpunctata L.), ��������� ����� (Sirex juvensis L.) �
������� ���������. ��-��������, ��� ������� �
���������� �������������� ������������� �
������������� ��� � ��������� ������� �
��������� ����� ��������, ��� ������������,
���������, ����������� �������������� ����
�����.
����� �������, �������, ��� ������� ������
�������������� ������� �� �������������
���������������� ����� (�������, ��������) �
�� ����-�������, �������� � ��� �� ��������� (��. Pterostichus
angustatus � Pt. vernalis, ��������� ������� ���������).
��� �� ����� �������� ����������� ������������
�����: ������� ������� (Agonum obscurum Hbst.), ���������
(Limoniidae), ��������� (Dolichopus sp.) � ��., ��� ������
��������� ��� ����� ������������ �����, ��� �
������� ����������� �������, �������� � �������
������ ���������� ������ � ������������
����������� �������� �����.
���������� ����� �������� �������
������������� ������������� ������ ����� ������
� ��������� �������������� (��� ������ �������
������������� ���������� ����� �����, ��� ����
���������� ������, �, ��������, ���� ������
������������ ����� ������ ����������, �� ��
��������� ������������ ��������). � ����� ������
����������� ��������� � ����������
������������� �� ���������� ��������� (0,96-0,89) �
�����������, � ��������� ���������� � �������
��������-��������� (0,83) (���. 2). ��� ������� �
����������� �������� ����������� �������������
�����, ����� ��� �������, �������, �������, �
���������� ������ ����� ����� ��� ����� (6,8
�����/�2) � ��������� ������. ����� �������,
� ���� ��������� ����������� ������������ ��
����������� ���������� ����� ����������� �����,
���, ��������, ����� ��������� �� ������
������������������ � ����� ���������� ��������
� �������� ����������.
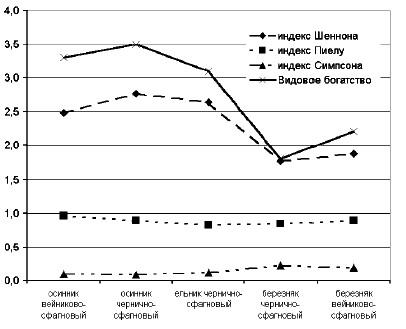
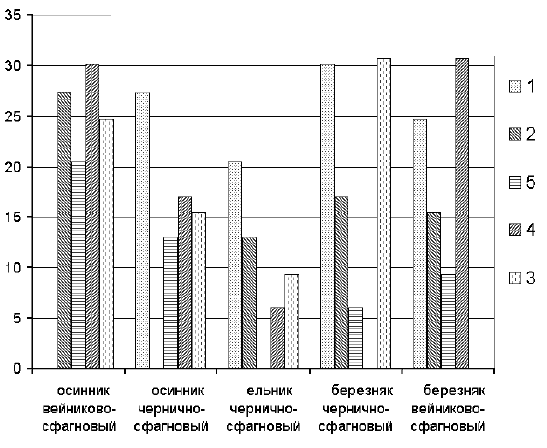
���. 2. �������������� �������
������ �������� ��������� ������������� �����
����� � ��������� �� ������ �����. ��� �������
���������� ������� ������������ �������
��������� ����������� � ��������
��������-���������, � ����������� �
���������-����������. � ���� ��������� � �
������� ������� ��������� ����, ��� � ����������.
������ ������� ���������� ����� �������
������������ ���������. �� ���������� ���
�������������� � ����� ����� � ���������
�������������� � ������� �������������
������������� ������ ����� ����. ��������
�������� �������� ������������ � ������
�������������� ��������� �������� ��������
���������. ��� ������� � ���, ��� ���������
������������ ������������ � �������� ����������
� ����� �����, � �� ���������� �������������.
�� �������� ������������ ��������� ������� ���
������� ���������� ����� (����������
����������� ���� � ��������� � ������� ��
��������� � ����������� ����� � ����������
�������� ��������� ���������), ��� � ����������
���������������. ����� �������� ��������� �
���������� ����������� ������ ����� (�������,
��������� ������� ���������). �����
������������, ��� ��� ������� �����������
���������� ���������� ���������, �� �������
����� �� �������� � ����� ������ �������� �
������ ������� ������� � ���� ������
"�������������" ��������. ��� �����
��������, ���������� �� �������������
���������������, ����� ��������� �
�������������� ������� �������� ����� ����� (����������
Malthinus sp. � Maltodes sp. � ������ �
������������������ ��������� � ��������� � ��.),
��� ����� ��� ��������� � ����������, ��� � ���
(��. Pterostichus vernalis Pz. � Pterostichus strenuus Pz.), � ���
�� ������������ � ������ ����������� Carabus marginalis
F. � Aphanus quadratus Fabr.
������ �������� ���������� "������������"
������������ �����, ��� ��� ��� �������� ���
������, ��� ������ ���������� ����������. ���
���������� ������������ ���������� � �������
��������-����������, �������� ���������� ��
�������� ��������-��������� (0,225), ���, � �����,
������� �� ���������� ����� ����������� �����.
������������� ���������� ��������
������������� �������� ��������� ���������
����������� ��������. ������ ������� ��������
�������� ����� ��������� �� �������� ������� �
��������� �� ������, ��� ��� � ������ ������
�������� ����������� ������ �� ������� �������
��������, � �� ������
������ ��������� ���� �� ����������� ����������
����� � ������� �� �������. � �������
������������ �.�. ���������� �� ����� �������
����� ������������� ��������, ������������ ���
���������� ����������.
�������� �� �������� ������� ������������ ����
�� ������ ������������ �������. ��� �������
���������� ����������� (����. 2, ���. 3) ��������
����� ����� �������� �� ������� ���������� �
�������� ���������.
������� 2. �������������� ������� �
������������ ��������
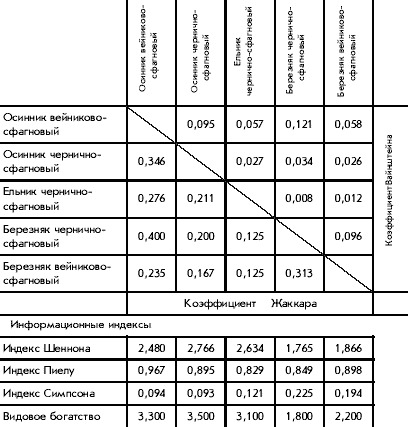
���. 3. ����������� �������� ���������
������.
1 � ������� ���������-���������; 2 � �������
��������-���������; 3 � ��������
���������-���������; 4 � ��������
��������-���������; 5 � ������
��������-���������
� �������� ��� ������, �������� ��������������
����, ������� ��� ���� �����������
�������������. ����� ������� ��������, ��� ���
������� ���������� �������� ������ ���� ��
�����, ��� ���� �������������� ���������. ��
������ ����� ����: �������� ��������-��������� �
�������� ���������-��������� � �������
���������-��������� � �������
��������-���������. �������� ���� ��������
�������� �� ���� ����� � ���������, ��������� �
������� �������� ��������� �
�������-��������������� �����, ����������� ���
������ ����� � ����������� �� ������ � ������. �
�������-���������� ����� ������ ��� ���������,
��� ��� ���, ��-��������, ����������� ��������
������ �� ��������� ������������ � ���������.
������� �������� ���������� � ���������
����������.
����������� �������� ��������� ������ � ����
������� �������� ����� ��������� ��
�������������� ������������. �������� �� ������
� ����� ��������� ����������� ������� ��������,
������������� ����, � �� �� ����� ������������
����� ����� ������ ������� �������������
�������� ������. ��� �������� ����������� �����
������� ��������� ������ (����� ����� ��������
��������-���������, � ������ � ������
��������-���������). ������� ���������
����������� ���� � ��������� ����������, ��� ���
� ��� ����� ��������� ���� �����, ���������� ��
���������� ������. � ���� �����, �������
��������� ����������� ����� � ���������
����������, ��� � �������. ��������, �� ��� ������
������������ �������� ���������� �����,
�������������� ������ ������� ��������
�������-��������������� � �������� ������. ���
������������ ���� ���������� ����������
����������� ������� � ��������������. ���������
�������� �������-���������� � ����������
����������� ������� ���� �� ������� ������
�����, ����� ��� ���� ���� ��������, � ������ ����
���������.
��� ������ ����� �������� ��������������
�������� ������������ �������� ���������� ���
�������� ���������-���������� � ���������
��������-���������� (0,121) (���. 4), ��� �������
����� ��������� ��������� ��������� �� ��������
�������������.
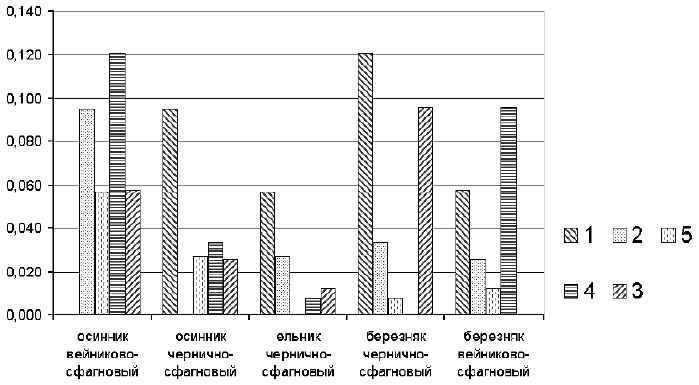
���. 4. ����������� ����������.
1 � ������� ���������-���������; 2 � �������
��������-���������; 3 � ��������
���������-���������; 4 � ��������
��������-���������; 5 � ������
��������-���������
��� ��� ��� �������, � ������� �� ���� ������,
�������� ��������� �������, �������� ���������
������� ������� �� ������������ �� ���������.
�����, �� ���� ��������, ������������ ����:
�������� ��������-��������� � ��������
���������-��������� (0,096), �������
���������-��������� � �������
��������-��������� (0,095). �� ����� �������� �
��������� ����������, � ����� �������� �
��������� ��������� ����� ����� (0,058 � 0,034
��������������) ������� ������� ��������
�������� ������� �������-��������������� �����.
� ������� ��������-���������� �������� ������
��������, ���, �� ����� ��������������,
����������� ���� ���������� ����������� �
������������ ������������ ��������, � ������
������������, � ��� ��������� � �����������
��������� ���������, ��� �������� �� ������
������������� � �������.
������
������ ������������� ����� �� ��������,
������ �������� �������� �� ��������������
��������� ����� � ������������ ����������
��������� ��������������. ���, �� ���� ���������
� ������� �������������� ������� �
������������� ������ ����������, ��� ������� �
���, ����������� ��������� ����� �����
�������������� � ������������ ������.
� ���� ������ ���� �������� ����������
���������� ������ ����� � ���� ���������, ���
����� ������� ������� ��������� � ��������� �
�������, ��� � ����� ������ ������������
��������� (����,1975), ��������� �� ����������
����� ����� ��� ����������� � ������������
(����������) �������.
����� �������, ��� ������� ��������
������������ ���������� ����� ������������ �
��� ���������� �� ��������� ������ ������ �
�������������� �����������, ��� ������ � ������
������, ���������� � ���������� �������,
����������� ���� ��������� ������. ������ � ���,
���� ��������������� ����� ��������
������������ � ���������� �����������
�������������, ��� ��� � ���� ����� �������
��������� ��� ������, � ����� ������� ����������
� ������.
�������� ������ ��������� ��������� ������� ��
������� �������������� ������ ����
�������������� ������� ��������-����������.
������ ������� ����, ��-��������, �����������
��������� ��������������, �����������
����������� �� ���������� ��������. ���
����������� �������������, ���,
����������������, ��������� ���, ������������
����������� ���������� ����� � �����������
����� � ��������� ����������� �������������� �
��������� ���������� �����������. ��� ������ �
����� ������������ ���� �����������
�������������, �����, ������, ������������
���������� � ��������� �������� ��� ����,
������� ������ � ������-����������
���������������� ����� ������ ��������������
��� ��������� ���������.
����������
��������� �.�. � ��������� ������� ������
�������� ���������� // ����. ����., 1967. �. 46. �7. �.
981-986.
������� �.�. ���� ������� ���������
�������������� (���������) // ������
��������-������������� ������������. �.: �����.
1975. 282 �.
������ �.�. ��������� �������
(Coleopthera, Carabidae) ����������� ����� ����� ����� //
������������ ��� (������������� ������������).
�.: �����. 1986. �. 110-115
������ �.�. ������������ ��������� ��
��������. �.: �����������. 1972.
���� �. ������ ��������. �.: ���. 1975. 740 �.
�������������� �.�., ������������ �.�.
�������� ������������ ��������� ������ �
������� ������������� ����������� ��� ���� �
������� // ������� ����. ���. III. 1997. �. 6-9.
����������� �.�., ������� �.�., ���������� �.�.
�������������� ��������� ����������
����������� (�� ������� ��������-����������
�����������) // ������������ �������������
����������. �.: �����. 1983. �. 230-242.
������� �.�. ����� ��������� � ��������
���������� ���������������� �� ����� ������ //
����. ����. 1979. �. 58. �6. �. 834-845.
������ �.�, ����������� �.�. ���������
��������� ��������������� ��������-�����������
����������� ���� ���� // ������� ����. 1997. ���. III.
�. 14-17.
���������� �.�. ���� �����������
�������������� // ������ ��������-�������������
������������. �.: �����, 1975. 282 �.
������ �.�. �������� ����������������
�������������� ��������� �������������� �
������ �� ������� // ������
��������-������������� ������������. �.: �����.
1975. 282 �.
***
������ ������ ���� ������������ � ������� �������������� ������ ������������� �����
"������� ����"
������ ����������������� ������, ��������� � ������������� ������ ����, ����� ����������
� ������� ����������������� ������. ������ ������� ����������������� ������ (����������������� �������), ����������� �� ����� �����, ����� ����������
���.
 ������� ������
"�������� ����"
��������� "Parus"
������ ���������
������� ������
"�������� ����"
��������� "Parus"
������ ���������
|


 ������������ �������� (��� PC-Windows) ������������
������������ �������� (��� PC-Windows) ������������
