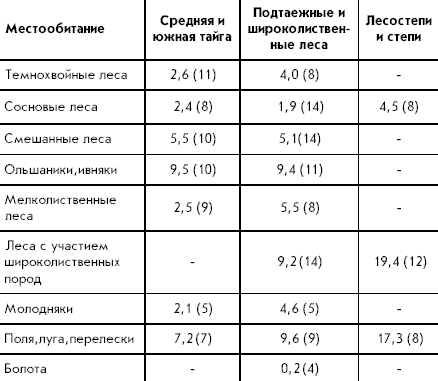
����������:
1) ������������� ����������������� ������ ��������� �������� �� ������ ������� ������������ � ��������� �� ����� � ������������ ���� (���������� �����������, ������� - �.�. ��������� );
2) ����� ����� �� ����� ��������������� �� ������������� ��������, ���������� � ������;
3) ������ ������ �������� ��������� � �� ����������� �������� ������ � ��������� ������� � �������� ������������ �� ���� www.ecosystema.ru
������ ������ ���� ������������ � ������� �������������� ������ ������������� �����
"������� ����"
�������������� �
������������� ������������� �������� ����� ������ �������� ���� ��������-�����������
������� (�� ������ ����������� ������ ������)
�.�. �������������� ,
�.�. ������
2002
�������� ����� �������� ���� �� ��������� �Parus� ���������� ��
���������� ������ � ������������ ����� � 1986 �. �
1992 �. � ���������� � ��� ���� �����������
��������� �������������� �������������� ����,
������������ �� �����������-���������. � ������
���� �������� ������ ���������� �������� � 20-35
������� �������, ������������� �� ���� ��������
����� �� ����� ������ ������, ��������������� �
����������� �����. ����� ����������, ��� �������,
3-5 ������ ����� �������������, � ����� ������� �
�������� ������������. � ������ ��������
��������� ��� ���������������� ����������
(��������������� ���������� ������������), ��� �
����������-�������� (� �������� �������� �
���������). � ��������� �Parus� ������������ �����
����������� ����� ���� ��� ����������� �������
������, � ���������� ������ �� ������� ���������
��� ����� ���� � ������ �����������, �� �����
��������� ����������� (������, 1967). �
�������������� �������������� ����� �� �����
����� ��������� ����������� ���� �� �����������,
� �������� �� ������� ������� � �������
�������������, ������������ ��� ������ ����� �
������ �� ��������� ����� ��������� ����������.
��������� ������ ������������� � ����� ������
����������� �������������� �����������
��������� ����������� � �������� �������� ��
���, � ����� ����������� � ���� ��������� ��������� ����������� ������
������ ���� ������ � ������������ ������ (���.
1-11, 1990-1999).
��������� ������ ��������� �������
����������������� ������������� �������� �����
�������� ������ ���� �� ������ �����������
������ �����������. � ������ ������������ ������
�� 60 �������� �������� � 398 ������, ������ �� ����
����� ������ ������ �� 1 �������� ������� �� 1
������ ������. ����� ������ ������� ������
��������� � ������� 1953-1957 �. (����� �.�.
������������� �� ��������). �������� �����
���������� ������� � 1986 �� 1999 �.
����� ����� ������� ������ ���������� �� ����
�������������� ������ ������������� ����� (134
����� �� 30 �������� �����������). ����� ����, �
���� ����� ������ 70 ������� � ��������� �����. �
�� ����� ���������������� ���������� �
����������-��������, ������������ ��������
������������� � ������������� ����������� ��
������ �����������, � ����� ���������
������������ ����� ��������� �������������
�����. ��� �������������� ������ �������� �
����� ������ ����������� ��������������
����������� ��������� ����������� � ��������
�������� �� ���. ������� ����� ������
������������ � ��������� ����������� ������
������-� (���. 1-11, 1990 � 1999). �� �� ����� �����
��������� ���� ���������� ���������� �������
�����, ������ ������ ������� ��������� 200
�������. ������ � ������ ������� ���������
���������� � ��������������� �������� ���������
����������� ������ ������-�. ���� ����������
������ �������� ��������������� ������, � �����
������������ ����� ��� �����������, �� �������
��� ������ ��������. ���� ������� �������
����������, � ����� ����������� ����� �� ���,
�������������� �� ��������, �� �������� ��������
�������������.
���� ���������� ������ ������� ������ �
����������� �������� ���� �����
������������������� ������� (� ������� �������
����� ���������� �����; ������� ������� � ������
������ ������������� �������� �� ������ � �����
����� ���������).
���������������� ���������� �
���������-�������� , ����������
������������� ��� ���������� ��������: ������
�.�. (���. �.������); ����� �.�. (���. �. �������);
������� �.�. (���������� ����������); ��������
�.�., �������� �.�. (���������� �������); ������ �.�.
(��������� ����������); ������� �.�. (����������
�-� ������������ ���.); ������ �.�. (����������,
������������, ���������, ���������� ���.);
������� �.�. (����������-������ ����������);
������ �.�. (����������� ����������); ������ �.�.,
������� �.�., �������� �.�. (������-��������
����������); ������� �.�. (��������� ����������);
������ �.�. (�������, ��������� ���.); ����� �.�.
(�������, ������������ ���.); �������� �.�.
(��������, ��������� ���.); ������� �.�. (��������,
������� ���.); ��������� �.�. (���������� ���.);
�������� �.�. (���������� ����������);
������������ �.�. (���������� ����������);
��������� �.�. (���������� ����������); ��������
�.�. (���������� � ����������� ���.); ������������
�.�., ������� �.�. (���������� ���.); ��������� �.�.
(����������-����������� ����������); ������� �.�.
(���. �. ������-���, ������ �. ����, ���. �.
���������); ��������� �.�. (���������� �������);
��������� �.�., �������� �.�. (�����������
����������); ������� �.�. (���������� ���.); ������
�.�. (����������� ���.); ����� �.�. (������������
���.); ������� �.�. (����������� ���.); ������� �.�.
(��������� ���.); ��������� �.�. (��������,
����������� ���.); ������ �.�. (���������� ���.);
������� �.�. (���. �. ����); �������� �.�. (���. �.
��������); ��� �.�. (���������� ���.); ������� �.�.
(��������� ����., ���. ���. ���������); ������ �.�.,
�������� �.�. (����������� ���.); ������������ �.�.
(��������, �����.); ���������� �.�. (�������,
���������� �����); �������� �.�. (��������,
��������� ���.); �������� �.�. (���������������
���.); ����� �.�. (�������, ��������); ����� �.�.
(�������� ��������� ���.); �������� �.�.
(���������� �-�); �������� �.�. (�������, ���������
���.); ������� �.�. (�������� ���.); ����� �.�.
(������������ ���.).
������������ �������� ������������� �
������������� ������� , ���������� �� ������
���������: ������� �.�. (��������� ����������,
���. �. ������ ������������ ���.); ��������� �.�.
(�������); ��������� �.�. (������������ ���.);
������ ����� ��� ������������ �������� �.�.
(����������� �������); ��� �. ������
(������������� ���.); ��������� �.�. (�������,
��������� ���.); ����� �.�. (�������, �����������
���.); ������ ����� (��������� ���.); ��������� �.�.
(���������� ���., �������);
��������� �.�. (�������� ���.); ������ �.�.
(����������� ���.); ��� �. ������ (�������������
���.); ��� �. �������� ���� (������������� ���.);
�������� �.�. (���. �. ���������); ��� �. �����
(��������); ���� ������������� �-�� �. ������
(��������� ���.); ���������� �.�. (�����������
���.); �������� �.�. (�������, ������������� ���.);
������� �.�. (����������� ���.).
������ ��������� ������������� ����� :
������� �.�., �������� �.�. � ������ (������� ���.);
������ ��� � ������� �.�. � ������ (���������,
���������� �����������); ������� �.�. ������ �.�.
��������� �.�. (�������, �������� ���.); ���� �
�������� �.�. � ������ (���������� ���.).
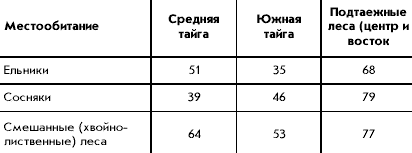
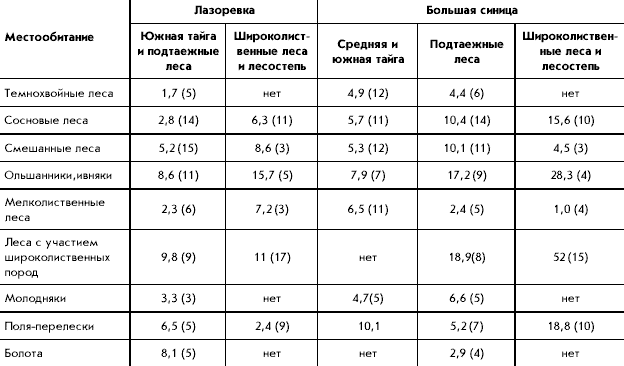
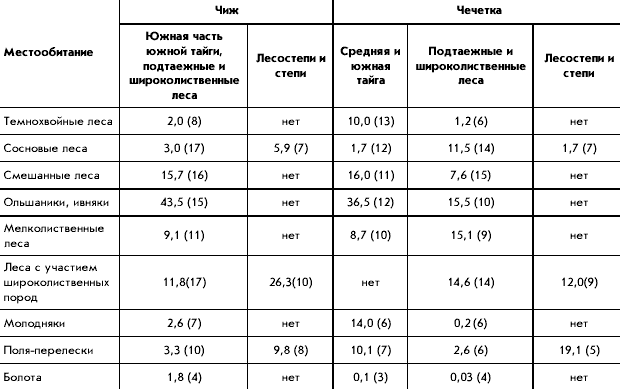
� ������ ������������ ������ �� 60 ��������
��������, �������������� �� �������� ���������
�������: ������ � 1 �����, �������� ����� � 6,
������� � 5, ����� � 9, ���������� ���� � 15,
���������������� � 5, ��������� � ����� � 13,
������ ���� ����� � 6. ����������� ���������� ��
�������� ������ ������� ���� �� ����� (�������
�������� ����� ������������ ������� � ��������)
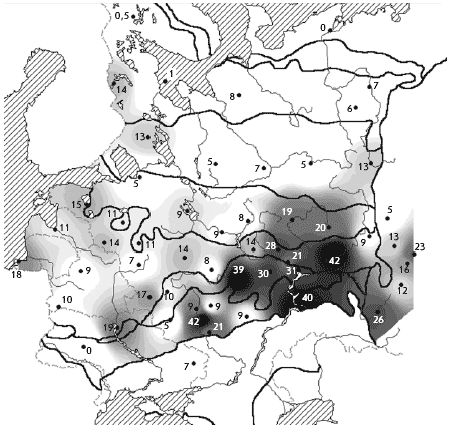
(���. 1). ��������� ������ ������ ����������, ���
������������� ��������� ����������� ��������
����� �� ���� � ���� � ������ ����� ������������
������� �������, �� ��������� � ���������
������� ����������� ����������, ��� ����
��������� � ������� 1-2 �������. ���� ����� ���
����� ����� ��������� ���������� ������ �� 3-5
���. �������� � ����� ������� ������� ��� ������
����� ��������� � ������� � ���. 1.
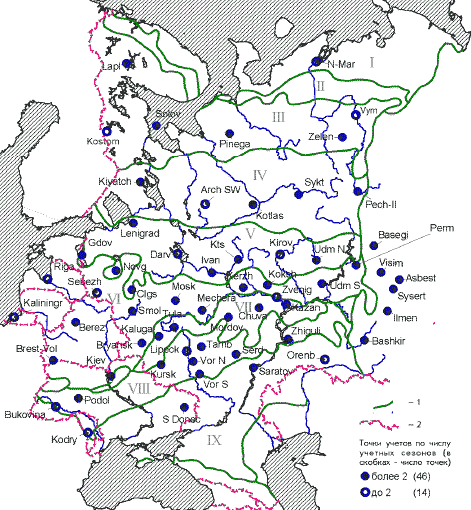
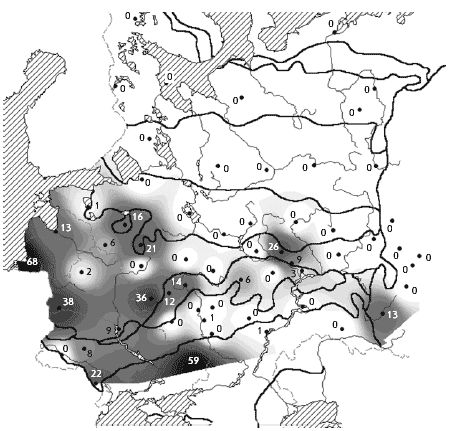
���. 1. ����� ������������ ��������
�������� ������ ������
1 � ��������������� �������; 2 � �������
��������� ���. ��������� ����: I � ������, II �
����������, III � �������� �����, IV � �������
�����, V � ����� �����, VI � ���������� ����, VII �
���������������� ����, VIII � ���������, IX � �����.
�������� ����� (� ������� � ����� �������
�������): N-Mar � ���. �. ������-��� (1); Lapl �
����������� �-� (6); Kostom � ������������� �-� (1); Solov
� ������� (4); Pinega � ��������� �-� (10); Zelen � ���.
���. ����������� (4); Vym � �. ���� (1); Kivatch � �-� �����
(12); Arch SW � ������������� ���. ���-�����, ���. �.
������ � ��������� (2); Kotlas � ���. �. ������ (4); Sykt �
���. �. ��������� (3); Pech-Il � ������-�������� �-� (5);
Gdov � ���. �. ���� (5); Lenigrad � ������������� ���. (6);
Novg � ������������ ��� (10); Clgs � ����������-������
�-� (14); Darv � ���������� �-� (2); Ivan � ���������� ���.
(7); Kts � ����������� ���������� ���� ��� (20); Kirov �
��������� ���. (2); Udm N � ����� �������� (4); Riga �
���. �. ���� (2); Kaliningr � ���. �. ����������� (1); Brest-Vol
� �������� � �������, ��������� � ��������� ���.
(9); Sebezh � ��������� ���., ���. �. ����� � ��������
(2); Berez � ��������, ����������� �-� (6); Kiev �
�������� (�������) (5); Smol � ���������� ���. (4); Bryansk
� �-� �������� ��� (13); Mosk � ���������� ���. (26);
Mechera � ������ (6); Kerzh � ���������� �-� (3); Koksh � �-�
������� ������� (4); Zvenig � �����-��, ���. ���.
��������� (1); Udm S � �������� �� (3); Perm � ��������
���., ���. �. ������ (6); Kaluga � �-� ��������� ������
(3); Tula � �������� ���. (3); Mordov � ���������� �-� (4);
Chuva � ������� (8); Kazan � ���. �. ������ (22); Bukovina �
�������, �������� (2); Podol � �������, ������� (9); Kodry
� �-� �����, ������� (1); Kursk � ���. �. ����� (5); Lipeck �
�������� ���. (1); Vor N � ����������� �-� �
����������� (13); Vor S � �� ����������� ���. (4); Tamb �
���������� ���. (4); Serd � ���. �. �������� (3); Zhiguli �
����������� �-� � ����������� (4); S Donec � �������,
��������� �-� � ����������� (3); Saratov � �����������
���. (4); Orenb � ������������ ���. (1); Basegi � �-� ������
(3); Visim � ��������� �-� (14); Asbest � ���. �. ������ (6);
Sysert � ���������� �-� ������������ ���. (9); Ilmen �
���������� �-� (12); Bashkir � ���������� �-� (11).
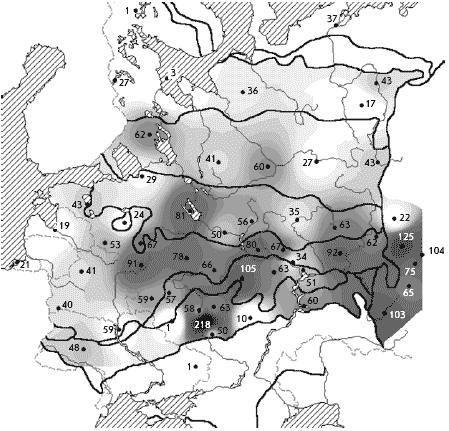
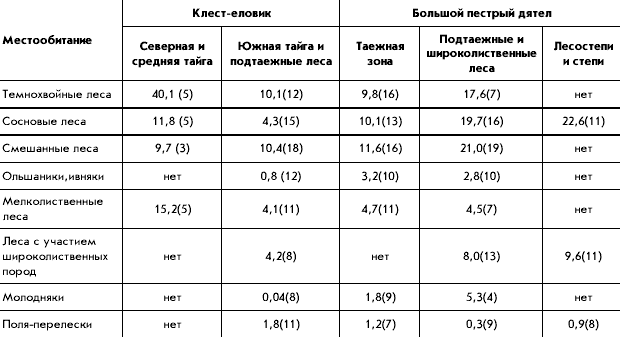
�������������� ������ ����������� ����������
�� ������, ������ � ������� �� ��������������
����� ��������. ���, ��� �����-����������
(������, �������� ������, ��������, ������������
�������, �����-������, ������� ������� �����)
����� ����������� � ������� �� ������� �
��������� �����, ��� ��������� ����� � �� ����
�����. ��� ����, ������� � ������� ��� ����������,
����� ������ ��������, ����������� ��� � ���� �
�����������. ������� ���� �� ���� ������� ��
���������� ��� �������������� ������� ����
���������. ����� ����������� ���� ���������� �
������� ��� MapInfo � �������������� ������
������������ (��������� �.�. ������� ).
��� ������������ �������������� �������������
�� ���������� ������ ��� ������� ���� �
����������� ��������� �������� �������, ��� ���
�� ��� �������, ��� ���������� ����������� � ���
�������� �������������� ���� ������������ ��
���� ���������� �������� ���������.
� �������� ������ ����������� ����
������������ ����� �������� ��������� �������.
�.�. �������� ��������� ���������� ���� ������;
�.�. �������������� ����������� ��������� �
����� ������.
�������������� � �������������
������������� �����
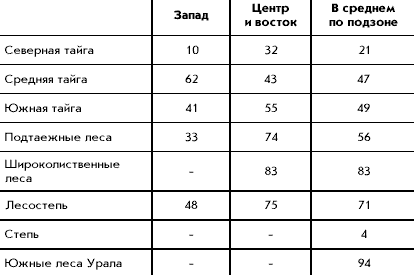
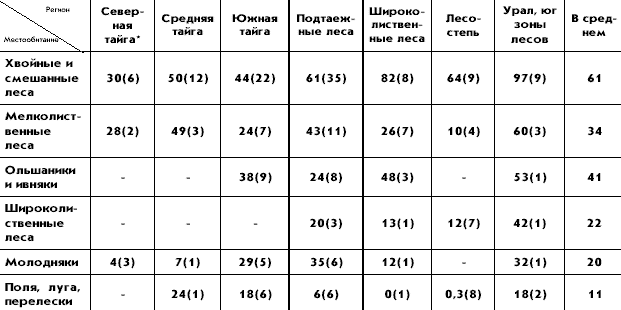
�������� ����� �������� ��������� �����
������������� ���������� ���������� 16 �����
����. � �� ����� 11 �����, �������� � ������
������� ����: ������, ����������� ������,
������������ ������, �������� ������, ��������,
������� ������, ���������, ������������ �������,
���������, ���������, ������; 4 � ���� ���������
� �����-������, ���, �������, �������; � ����
����� � ������� �������. ���� �� ���������
�������� ��������� ��������������� �
�������������� ������������� ��� ���� 16 �����,
���������� � ���������� ������� ����������
����� ������. ����� ����, �� �������� �������
�������� �� ������ ����� ����, ������� ��
�����������, ������� ����� ������� � ��
�������������� � ������������� �������������� �
���������� ��� ��������� ��� ���������������.
��� ������ ����� �� ���������� (� �������� ��
�Handbook of the Birds of Europe�, 1993; ��������� ���������
������� � �����).
������
������ � �������� �������� ��� ����� ����,
�������� � ����� ������������������ �������. ��
������� ����� ������� �������, ��� �������,
���������� �� ����������� ����� ������ ����. ���
���� ��� �� ������ �������������, �� � ����������
���������. ���������������� � ������������
�������� ������� ����� ���� ���������
������������� �� ��������� ��������� (��������,
��������������, ���������, 1988). ���, � ������
��������� ������� ������������ ��������� ��
������ ����� �������� � ��� ���� ��������
��������� ����� ����, ��� �� ���������� ���
������ ����� ����, �������� � ������ �������
����. ��������, ��� ������ � �����-�� �������
�������� �����-�������������, ������������
�������, �������������������
������������������� ������ �������� ����
(�������, 1987).
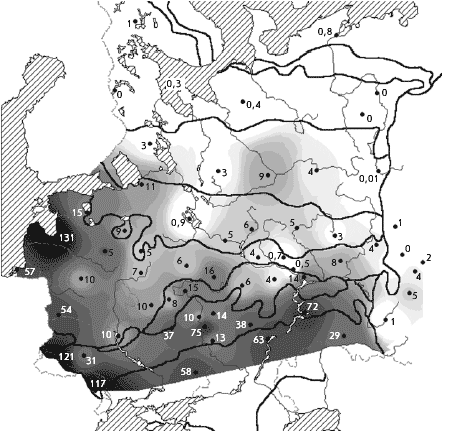
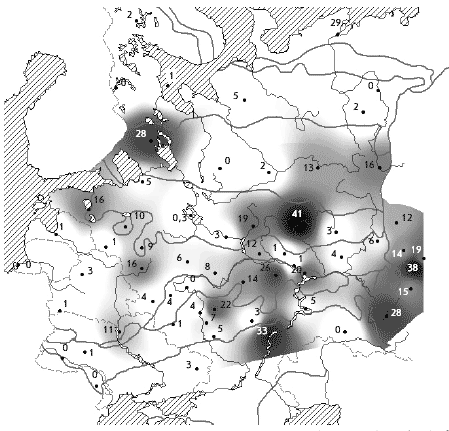
�� ������ ����������� ������ ������, � ��������
������������������� ������� � ����� �������
������������� ����� �� ���� ������� � ���������
����� � �� ���������� � �������� ����� ��
���������. ������������ ������� �� � ��������
����� ������������� �������, � ����� �������
���� ��� ����� ��� ����������� (���. 2).
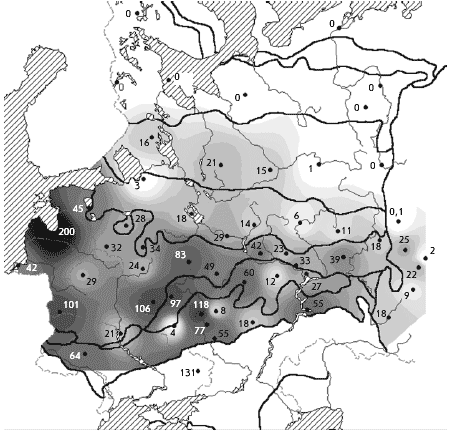
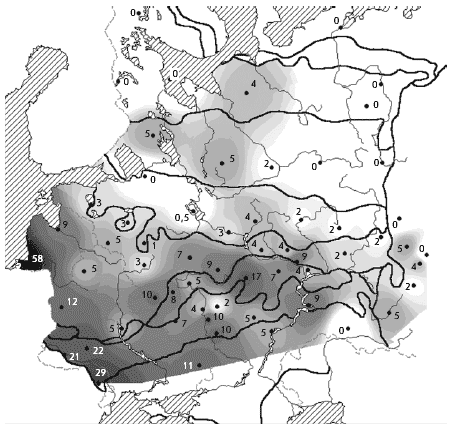
���. 2. ������. ������ ����������� �
������� � ��������� ����� (� �������), ������/��2
����� �� �����, ����� ��������, ��� �����������
��������, ������ � ������� �� ������� � ���������
�����, ������������� � ������ �� ��. ���, �
������� ���������������� ����� � � ��������� ���
� ������� �������� � 2 ���� ������, ��� � �������
�������� (����. 1).
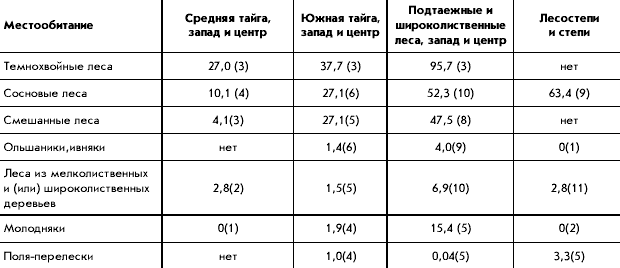
����. 1. ����������� ������� � ���������
�������� ����������� ������ � ������������
�����2 )
��� �������� �������� ������������, �� � ����
������ ����������� ����� ��������� ���������� �
������ �� ������. � �������� � ������� �����
����������� �������� �� ������ � �������
��������������� ���������� �����������
���������. ������ ����������� �� �������� �����
���� ��������� ��������� ����������, � �
����������� ����������� � ��� ������������� ��
�������� ������. � ����� �����, ���������� �����
� � ��������� �������� � �������� ����� ������ �
1,5-2 ���� ������, ��� � ������ � �� �������.
�������� ������� ����������� �������� �� ���
����� �����. ������ � ������� ����������� �
����������� ����������� (218 ��./��2 ) ����
������������, �.�. ���� ����� � ������� ������
������, ������������ �� ���������� ���
�����������.
����� �� �����, ����� �������� �����, ��� � ����
���������������� ����� � � ��������� ��������
����������� � ������ ������ �������� �������,
��� � �������� �������. ��� ����� ���� ������� �
������� ���������. ��-������, ����� ���������
������� ����� ����� ������ ����������� ���
����������� ���� � ����������� �� �� ��������� �
������� ������ ���������, ����������� �� � �����
��������� ������. ��-������,
������������ ����� ������� ����� � ���� ��������
������������ ��������� ���������, �������, �����
����� ������, ��� ���� ������� �����
��������������, ��� ������������ ����. �������,
����� ��������� ��������� ����������� �� �����,
����������� ��-�� ������ �������������
����������.
���������� ����������� ��������������
������������� �������. ���� �������� � �������,
�� ����� �������, ��� ������� ������
������������ �������� ������� � ��������� �����
(����. 2). � ���������� � �������, �
��������������� ����� �� � 1,5-2 ���� ������, �
���������������� � � 3 ���� ������, ��� � ����� �
�������� ������� �����. ������������� �������
����� � ������� �����, ����� � ����� �
�����������.
����. 2. ������������� �������������
������� � ������ �������� (������/��2 , �
������� - ����� ����)
������ ������������ �������� �������� �� ��
���� �������� ���������. ���, � �������� �
������� ����� ����������� �������� � ����������
����� ����������� ����� ��, ��� � ��������� �
�������, � ����� � ���� ����� � 1,5-2 ����, � �
�������� ���������������� ����� � ��������� � �
4-6 ���. � ����� ����� ������������� �������� ��
�������� �������� ���������, ����� ���
������������� �� ���� ������������� ��������. ��
������ � �������� ��������� � ������� �����. �
������������ � ��� �� ������� ���� ������
���������� �������� ����� ���������� ��������, �
����� ��������������� ���, ��� ���� �������
�������. ������� ����� ��������� ����������� �
�����-����������, ����� � � ���������� �����.
�������� ������������ ��������������
������������� ������� � ����������� �� ��������.
���������� ���������� ��� ��������� �
���������������� �����, ������� �������������
������� ��������� ������������� � ������ ��
������. ���, � �������� ������� ���������� �����
(�������, �������� � ������������ ���.) �
���������������� ���� �������� 9 ������/�� 2 ,
� ����������� � 13 (���������� ��������� ���) � 37
(��������� ����������, ���������� ��������
�������), �� ����� � 42 (��������, ����������
�������-���).
��� �������� �������� ����������� �������� �
������ ����� ������� � ��������� �����, ��
����������� �� ������� �� ��� ���� ��������, �.�.
� ����� �� ��� ��� ��� ���� ��� ��������������
����������� ������������. �� ��������� ������
����� �������, ��� �������� ��� �������� (����. 3).
����. 3. ����������� ������� � ������
����� ����� � �������� ������� ����� (������/��2 ,
� ������� - ����� ����)
� ������� ���� ������� ��������� ����
����������� � ��������� �����, ��� � �����
�������, � ���������� ����� ��� �������� ��
��������. ���� ��������, ��� ������� ��
����������� ������� ����� ��������� �� ������
���������, � ������� ��� ������������. ������ �
����������� �������� ���� � ������ ����� ���� �
����� � ��� �� �������� ��������� �������. ��� ��
�����, ��������, ��� � ���������� ������������
�������� ����������� �������� (��� � ���������
����������) ������� ������, ��� �� ������� �
����� ����� ��������� ����. � �������������
������������ ����� �������� ����� �������
������������ ������ ����������� �������� � ���
�� ������ � � ������� ��������.
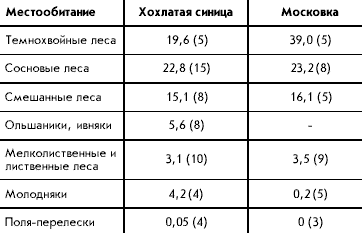
�������� ������
�������� ������ � �������� ���-��������.
��� ������������ �������� � ������� � ���������
����� � �������� ���� � ������ ����� � ���.
�������� ������ �� ������������� ������������
��������. �������� ����� ������� �
����������������� � ������� �������� ����; ��
����� ������� �������� �������� ���� �����
���������� �� ������������ ����� �� ������ ����
������� ������ ����, ��� ���������� ���������
(������, 1983). ������� ���� ����� �������� ������
����������� � ������� ��������� ���� ������ ���
����������, ���� � 3-4 ����� � ����.
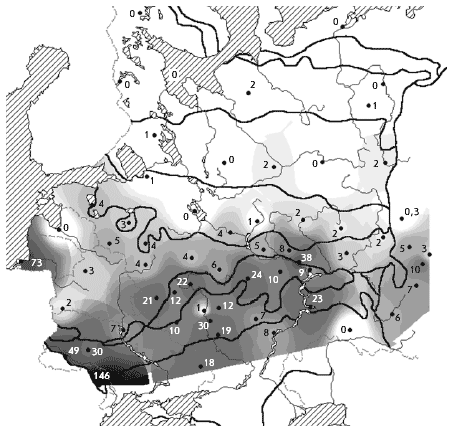
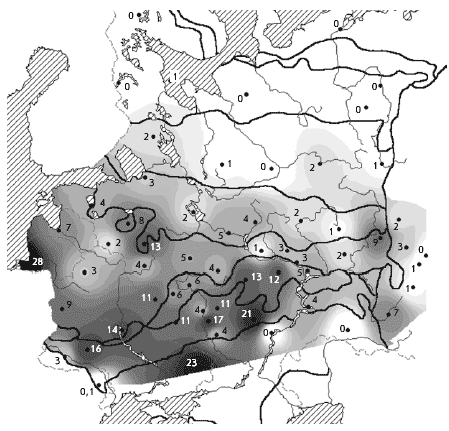
�� ������ ����������� ������ ������ ��������
������ �� ����������� � �������� ����� �
��������� �������� ������� ����� (���. 3).
����������� ����������� ��� � � ���������, ���
���� � �������� ����� ��������� ��������. �����
��� � ��������� ���������� ���������� ���� �
������ ����� ������������� �������������.
��-��������, ��� ������� � �� � �������� �������
������� ����� � ������������� � ���������.
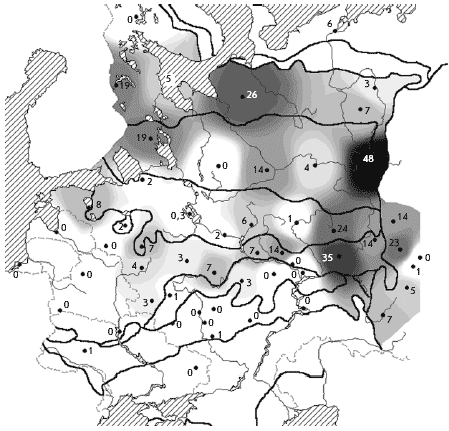
���. 3. ����������. ������ ����������� �
������� � ��������� ����� (� �������), ������/��2
��������������� �������� ������ � �������
���������. ������� ���������� �� ������ � �����
����� ���������� ���� �� ����� ������
������������� ����������, � ���������� �
���������������� ����� ������� �� ������ ��
������������� ������� � �������. ������, �
�������� �������������� �������, ��� ��������
������ ������ �����, ������� ����������� ��
���������� ������ �� �� ���� ��������. ���
���������� ���� ������������ ����������, ��� ��
������� ����������� �������� ���� �
��������������� ��������� ���� �������, �
�������� ���������� � �������������
������������ � ��������������� �����. ���, �
���������, ����� ������ ��� ��������� ��
����������� � ������ ������� �����������, �
����� ����������� � ����������� ��������� ���
(��������� ��������, ����������� ������� ��
������ � ������) � � ���������� ��������
(�������� ��������, ������� � ������ � ��������,
����������� ���������� ���� � �������������).
���������, ��� ��� ������� ����������� ��������
������ � �������� ������ ��������� �������
������������ �� �������������� � ��������, ���
�������, �� ��������������, ��� ���������� �����
����������� � � ������ ����� � �������� �������
����� (����. 4). �� ������ ������������ �����
�������� ������ � �� ����� ����� (���������, 1986).
����. 4. ������������� �������������
�������� � �������� ������ � ������� �� ������ �
������� ������������ (������/��2 , � �������
- ����� ����)
��������
��������, ��� ��, ��� � �������� ������ �
�������� ���-��������. ��, � ������� ��
����������� ����, �������� ������� ���������, �,
����� ����, ��� ��� ���������� �������������
������� �� ������-��������� ���� ������ � ������,
��������� �� ������������ ���.
�������� �������� ����� ��������� �� ����
�������� � �������� �������������� �������,
������, �� ������� ����� ���������� ���
���������������. ���, � �������� ����� ��������
�����, �� ������� ����� ��������� ������ �
������. ������� ����������� �������� ����������
������ ��� ��������� ����� ������ ����������,
���������������� ����� � ��������� � ��� �����
����� ����� (���. 4). �� �� �������� ������������
�������� ������ ����������� �������� � ����
����������� �������� � �������� � �����������
������ ������������� ����������. ��������, �
���� ������� �������� ��������� ��� �������
��������� �� ����� ������� ������� � �� �������
����� �� ��������.
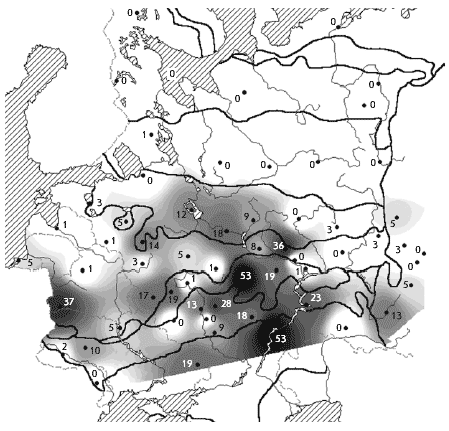
���. 4. ��������. ������ ����������� �
������� � ��������� ����� (� �������), ������/��2
������ �������������� ������������� ��������
� ���������� � ������� ������������ ����������,
��� ��� ������ ����� ������������ ������������
�����; � �������� � ��������� ����� �� � 1,5-2 ����
������, � ���������� �����, ���������� �
�����-���������� ��� �������� (����. 4).
������������ �������
������������ ������� � ���, ����� �����
��������� � ����. ������� ��� ����, �������� ���,
����������� ��������� ������������� �
������������ �� ������ ������ � ���� ���. ������
������� ���������� ��� � ��� ��������� ������.
�������� ������� ���������. ������ � �����
������ ������� ������� �������������
������������ ��������� �������� ��
������������� ���������� ������� ������ � ��-��
������ ��������, � ����� ��-�� ����, ���, � �������
�� ����������� �������� ����, ��� �� ����������
��� ������� ����� � ���������� �������.
�������� �� ����� ����������� �������� �� ���,
��� ���������� ����� ������ ������� �� �������
��������������� ������ �����. ��
������������������� ������� �������� �����
����������� �����, ����� �������� �����. ���, ���
��� ��� � � �����, ��������� � � �������
���������������� ����� � ��� � �������
����������� ������ � �������� (���. 4).
����������� ��������� � ������ �� �� (�� �������
����� �� ������� �������) ���������� ����� � 10
���; � ����������� ������ ��� ������� ������
����� � � ������� �� �����. ����� ��������
�������������� ������������ ����������� ����
������, � ������ �������, � �������������
�������. ����� ��������, ��� ��������� ��������
����������� � ������� ��������, ��� �������, ��
������ �������� ������� ������������ �� ���
�������, ����, ������, ������������ ��� �������
����������� � ���������.
���. 4. �������. ������ ����������� �
������� � ��������� ����� (� �������), ������/��2
���������� ����� ������ ����������� ���������,
����� ��������, ��� �� ������ ������ ����� �����
� ���������� ����� (����� ����� ������� ��������
�����) ���������� ������ �������� �����������
������, ��� ����� ���� �� �������, ������ �� �����
��������� ��������� ����������� � ���-������.
��� �������� ���������� ����������� ����������
�������������, ���������, ������������, ��������
� ���������� ������� � �������� ����� ��������.
��������� �����, ������������ ������
����������� �������� ����� ��������� �����
����� ������������� �������: ��������������
������� �������, ������ ��������, ������
����������� ��������, �������� ����� �
����������� ����������� ������ ��� ���������
����� � ������ �� ��������. ����� ������� ���
��������� �������������� ����� ��� ��, ��� �
������.
� ����� ������������� �� ��������������
�������� ����� �������������� ������� �
��������� ����� � �������� ���������� ����,
��������� � ����-��������� (����. 5).
����. 5. ������������� �������������
�������� � ������ �������� (������/��2 , �
������� - ����� ����)
����������� ������
����������� ������ � ���, ������������ �
����� ��������������� ����� ������������� �
������������� �����. �� ����� ������, � ������
����� ����������� ������ ������������� �
����������� �������� ������� �������� �����
(����� ���������� �������� � ��������������
�����������). ��� ���� ���������� ��������� �
����� ����������� � ������ �� ������ (���. 5).
������� ����������� �������� ����� � ���� �����
�� ������� ������� � � ����������� ���� ���
������-�����. � ������� ����� � ����������� �����
������ ����� ��� ������; ������������ �������
����������� �������� ������ � ���������� � �
������-�������� �����������. ����� �����
���������� ����� ����������� ������ ����������
� ��� � �� ������ ���������� ����� (��������
����� �����, ��� ��� �������� � ���������
���������� � ������������ �������). � ������ ���
����������� ��������� ����������� ������ �
�������� � �� ����������� ������� ������� ����
��. �.�. ���������, � ������������ ������
����������� ������� (������� ����� �����). ������
����� ��� ����� �� ��������, ���� ������� ������
����������� � ������� ����� 15 ������ �������.
���. 5. ����������� ������. ������
����������� � ������� � ��������� ����� (�
�������), ������/��2
� �������� ������� ������� �����������
����������� ������ ������������� � ���������
�������� ����� ���������� ������� (����. 6).
������� ������ ���������, ������, �� ����� ���
��� ������� ��������, ���� � ������������ �����
�� ��������� � ���������, ���������� �
���������������� �� ����������� ������� ����. �
�� �� �����, ������������ ����� ��������������
����, ���������� � ������� ����������� ������
��������.
����. 6. ������������� �������������
����������� ������ � �������� ����� (������/��2 ,
� ������� - ����� ����)
������������ ������
������������ ������ � ��������
������������������� ������� ����������� �
�������� � ���������� ����� �� ���-������. ��-��
�������� �������� � �������� ����������
������������� ����� ���������� ���� ���, � �����
� ��� � ������ ��� ����������� � �������������
��������� ����������. ������, ����������
���������� � ���� ����� � ��� ������, ��� ��
���������� ������, ����� ������, ��� ��� ���� ������ �����
������ ���������� �� ������. �� ����� ��������
������������ ������ ����� ���������� �� �������
����� ������. ��� �� ������� � ��������� �
����������, � ������� �� �������, � �������� �
���������������� � �������� �����, ��� �
��������� ���� � ������������ ������� ���������
�� ������� ��������.
�� ����� ������, �������, ��� ������������
������ �������������, ����� � �������� ��
�������� ����� ����� ������� ����� ����� ��
������� ����� � ������ �� ��, � � ��������
����������� � � ������ �� ����� ������-�����
(���. 6).
���. 6. ������������ ������. ������
����������� � ����� ������� ���� (� �������),
������/��2
����� ��� �������� � ������ ������� ���������,
���������������� ���� � ���������� ���� �
�������� ���������������� �����. ����
����������� � ��������������� �����, �������� �
�����-����������; ���������� � ������������
����� �������� (����. 7). ��������� ������
����������� � �������� ���������������� ����� �
��������� � � ����� ����� ����� (� ��������), ��,
��� �������, ����� ��� ������. ���������� �������
��������� ����������� ������������ ������ �
��������� �������� ����������� ��������
������� (�����-��). �����, ������, ���������� �
������������ ������� ������������� ���������
������.
����. 7. ������������� �������������
������������ ������ � ���������� (������/��2 ,
� ������� - ����� ����)
� �������� ������� ������� �����������
������������ ������ ����������� �������, ��� ���
������������ ��� �����������. ��������������,
���, ��� � ��� ������� ���� �����, �������� �
�������� ������� ����� ����� � �������� ������,
��� ������������ ������ ������������ ��������
����������� ����������� ��������� ���������.
���, ��� ��� ����������� � ������������
�����������, �� ������� ���� � �����������
������ ������� ������� �����������, �����������
���� ����������� ������� ��������� �������� �
���������������� �����, ���� �����������
����������. � ��� ������, ��� ������� ����� �����
�������� ��� �����������, ������ ��� ��� �����
����. ����� ����������, ��� �����������������
����, ��� ����������� ��������� ������,
��-��������, ������������. ���, ������ ��� �
�������� ����� �����������, ������, �����������
�����������. ��� ��� ���� �� � � ��������, ���
���� ������ ����������� ��������������� ���
��������� ���� � �������� ����������������
�����. � �� �� �����, ������� ��������
����������������� ���� ��� ��������� ����������
��� ��������� ����� � �������� ����������������
����� � ����������� ��� ������������
������������ ����������� ������, ��� �������,
����������. � �� �� �����, ��������� ���������
�����, ������, ����� ���� �������� ���������� �
��� ������. ���, �� ����������� ��� �
���������������� ����� ����������� ������� (�
����������� � ��������� ������������), ����
����� ������������� ��� ��������������
������������� ��� ������������ ��������� �
���������� ������� ������������. � �� �� �����,
�� ��������� �����, ��� ������ ���� � �����������
��������� ��������� ����� ������ �� �����������,
������ � ���� �� ������������ �� �����������
�����.
���������
�������� ���� ����������� � ����������� ��
��������� ������������� � ������������ � �����
����� �� ������ �������� ������ ��������.
���������� �� �������� ���������� ����������,
��� ���� ����� ��� �������� �� �������, ��
�������� ����� �����, �����, ���. ����������
������ ������������� � ������� ���������. ���
�������� �������� � ���������� ��������, ���
�������� �� ���������� ������� ������.
�� ����� ������, � ��������
������������������� ������� ���������� � ������
������ ������������� �� ����� ����� �������
����� ����� �� ���������, � ������� �������
����������� ���������� � ��� ����� ����� �����
(���. 7). � ������� ����� � � ������� ����� ��� �
������� ������, � �������� � �����. ��������
������������ ����������� �������������� �����;
������ � ������� ���������� ����� ����� ��������
����� ������ ����������� � �������� �������.
����� ���������� � ������� ������, ����� ��� �
������ � �� ������� ������� � �������������.
����� �� �����, ����� ��������, ��� �� ��������
�������� �������� ����������� ����������� �����
������� ����������. ������, ������� �����-����
�������������� � ���� ��������� ��� ���� ��
�������. ��������, ��� �������� ������
����������� ����������� � �������� ������������
��������� ����������� ���� �� �����.
���. 7. ���������. ������ ����������� �
����� ������� ���� (� �������), ������/��2
������ �������������� �������������
���������� ��������� ������� ��� ����������
���������� �����. �� ����� ������ ������ �
�������������� �������� �� �������� ������
������������ ���� � ����-��������� (����. 7). �
������� �� ������ �������� ��� �����������
������������� � ��������� ����, ��������� �
������. � ��������� ���������� �����, �������� �
���������� �� ����������� �������� � �������
���� ����.
���������
��� � ���������, ��������� ������������� �
������������ � ����� �� ������ ������ ����������
��������, �� ��������������� �������� ��������,
����������� ���������������� ������, ����� �
���. �������� ����� ��������� ����� ��������
����� �� ������������ �������, ��� ������������
������ ��������� � ��������� �������� ������
�������� ���������.
� ����� �������������� ���������������
��������� ���� �������� � ���-������
������������������� �������. ������������� ���
������ � �������� � ����������� �������� �������
���������������� �����, ��������� � �����, �
����� �� ����� ���-������ ������� ���������
����� (���. 8). � ����� ����� ��������� � �������
������, � ������� � �����, � �������� ��
�����������. �� ���� �������� ���������� �����
��������� � �������� ����������� � ������ ��
������.
���. 8. ���������. ������ ����������� �
����� ������� ���� (� �������), ������/��2
�� ����� ������ ������ � ��������������
������������� ��������� �������� ��������� �
������, � ����� ���������� ���� � ��������
���������������� �������� (����. 8). � ���������
����� ����������� ���� � 1,5-2 ����, �
��������������� � �������� � � 2-3 ����. ��������
������������ ��� ��������� ������������ ����.
����. 7. ������������� �������������
��������� � ������� ������ (������/��2 , �
������� - ����� ����)
������� ������
������� ������ � ����������� ������
��������� ���������� �������. � ������� �
�������� ����� ������� ������ ���� �����
������������� ��� ���� ������ �������������
(����� 100 ������ �� 1 ��2 ). � ��������� ��
�������� ��� ��������� ��������� � ����������, �
�������� ���������������� �����, ���� �������
������ � �������� ����� ����� � ������������ �
������������ ������� ������� ����.
�������� �� �������������� ������� ������ �
������ ����� � ����� ��������, �� ����
������������������� ������� �� ����� ���������
� � ������ ��������������. ��� ���� �����������
����� ������������� � ������-������� ��
���-����� (���. 9). � �������, ������� ����������
������ ���������� ��� ����� ������, ������� �
������� ���������������� �����. ������, � �����
�������� ������� ����� ������� ������
������������� � � ���������� �����, � � �����
�����. �������� ����������� � �������
�������������� �� ���� ������ ��������.
���. 9. ������� ������. ������
����������� � ������� � ��������� ����� (�
�������), ������/��2
���� �� ����� � ������ �������������� �������
����� � ���������� �������, �� �������������
������������� � ���������� ����� � ����� ������
�� ������������� ���������. ���� ����� ���
����������� � ���������� ����� � ��������
���������������� �����, ���������� � �������
(����. 8). �� �������� ����� �������� � ���������
�����, ����� � ����� � �����������. ��������
������������ ��� ��� ������������ �
��������������� ����. ��� �������� �������
������, ��, ������������ �������������
������������� ������� �����, ����� ������ �
������, ��� ����� �� ����������� ��� �����, ���
���������. ������ �����, �� ����������� �
�������� �������� ������� �� ������� �� �������
����������� �������������, ������� �� ��������
������������� �����.
���������
���������� ����� �������� �������� � ���
����������������� � ������������ �
�������������� �� ������� �������� �
����������� �����. ����� �� �������, ��������
���������� � �������� �������� ��������������.
�� �� ������ �� ���������� � �������� ����
������, � ������ ������� � ������� ������
��������� ��������: ������, ������ �����, ������
���� � ����, ������� �������� �������������,
������� � ������� ����. ������������ �����
��������� ���������, ������, ��������������
������� ���, ������� �������� ������. ����
�������� ���������� ������� �������� � ��
��������� �� ��� ������ ������ ������ ����, �����
���������� ���� ��� 1-2 �������. ������� � �������
�������� ���� ����� ���������� ������ ������
����-������� ���������.
�������������� ������������� �������� ������
�� ������������� ��������� � ������� ������. ��
����������� � ������� ���������������� �����,
����� � ���������, ������ ������������
���������� ������ ���������� � ����� ��������
������ ������� � ���, ��� ���� ����������������
���� � �������� ����� � ���� (���. 10). ����, ��� ���
��������������, ���������� ����������� ��������
� ����� ����������� ������� (146 ������/��2 )
������ �����, �������. � ���������� ����� � �����
����� �������� � ������� ������, � ������� �����
� �����, � �������� �� ���� ����� �� ����������. �
������� �� ������ ����� � ����������
���������������� ����� � �������� �����������
�������� � ������� � ����������� ������, �����
����� ������� ����������� �� �������� ������
���������� ����� � ���������, ����������� ��
��������. ��� ��, ��� � ��������, � ��������
���������� ������� ���������� ����������� ��
������-������ ������� ���������� �����, �� � ���
��� �������, ������������ ������.
���. 10. ���������. ������ ����������� �
����� ������� ���� (� �������), ������/��2
������������� ������������� �������� �
�������� � ����� ������ ��� ����������� ������
��������� �����������. � ����� ����� �
���������� ����� �������� ����� � �������� �
�������� � ��������� �����, ���� ���, ��� ����
������� ����������������� ����, �������� ��
����� ������� (����. 9). � ����� �������� ��������
������������� ��������� � ����������������
���� � ���������. � ���������, �������� ����� �
���������� ����� �� ������������ ��������
��������������� �������� ����������� � 1,5-2 ����
������. ������������, ��������������� ����,
��������� � ����-��������� ����� ��������.
����. 9. ������������� �������������
�������� � ������ (������/��2 , � ������� -
����� ����)
������������� ������������� ���������
���������� ���� ������� � �������������
��������� ���� �������������� ����� ��������. ��
������ ����������� ���� � ������� �����,
��������� ��� �������� � ���, �����, ������,
����� �����. ������������ ������
���������������� �������������� ��� �
������-������������ ����� � �������� ��������
������������������� ������� � �����. ���
���������� ������������ ������� ������ ��������
� �������� � ��������� ����� � ������ � �
����������. � ��� �� ����� ����� ����� ��������
������, ���� � ������� ������������, � �� �������.
������� ����������� ��������� � ����������
����� �������������. � ���������������� �����
�������� � ������������ �����, ��� �������,
����������. ��� ���� �������������� ���
���������, ����� ����, � �� ���� ������ ��������
������. ����� �������, � �������� ����� ������
����� �������� ���� ����� ����������� � ����� �
�������� ����� � ���������������� ��������. �
��� �� ������������� ������� ��������� ��
���������������� � ������������ ����.
������������ � ��������������� ����,
������������, ��������� � ����-��������� ���
��������� ���� ��������������.
������
������, ��� � ���������, ��������� �
�����-������������. ��, � ������� �� ���������,
������ ����� �������� �� �������� � �����
�����-������������ �����, � �� ���������� � ����
������� ������ ��������. ������������ �����
����� ��� ��, ��� � ���������, ��������������
������� ��� �, ���� �����, �������� � �������
������ �������� ���� ������ ��� ����������.
������ ����������� � ������ ����� �� ����
������, ����������� � ������� ����������
������������������� �������. �� �����������, �
��������� � ������� ��������� ������, ����
���������� �������������� ���������� �
����������� �� ��������� 20 ������ �� 1 ��2
(���. 11). �������� ������� ���������� ������
���������� � ����� ���������������� �������,
��������� � �����, � ����� �
����� �������� ����� ������� ���������� �����. �
�������� ���� ������ ����� ��������, ���� � �����
����������, ���������� ����������� � ������. ��
������-������ ������� ���������� ����� � ������
��� ��, ��� � �������� � ��������, ���������� ����
���������� �����������, ���� � ����� ����������.
� ����� ����� ������ � ������� ������, � ������� �
�������� ����� ���� ����� �����.
���. 11. ������. ������ ����������� �
����� ������� ���� (� �������), ������/��2
������ �������������, ���������� ��������,
���������� �����; � ���� ������ ��� ���������
���� � �������� ������� �����, ��� � ����,
���������� ���������������� ���� �������� (����.
9). �������� ��������� ������, ��� �� ��� �
��������, ��������, ��-��������, ��-��
������������ � ��� ����� ����� � ������� �����. �
����� �������� �������� ���� ���������������
������������� ������������ �����������. �� ����
�������� ����� ��������, ���� � ����� ����������,
������������ �������� ����� � �������� �������
�����. ��������������� �����, ���������� �
�����-���������� ������ �������� �����.
�����-������
������-������� ������������� ����
���������� ����������������� � ������� ��������
���. ��� � ��� ����������� ��������� �� ��������
����� ������, � ����������, � �������� ������ �
������������� ����� �����������, � ����������
���������� �������������. ��� �������
���������� ����� ������ ��������� �����������: �
��� ������ ����� ��� �� ����� ���� �����������, �
����������� ���� � ����� ��� �������������.
��������, ��� ������� ������ ����� ��� � �����
������������������� ������� �����������, �
�������, ��� � 4 ����. � ����� � ����, ����� �������
����������������� ����������� ������, ������
����� � ����� �������� ����� ����������
��������� � ������� �� ����� 8 ������� (�����
��������� �� ������� ���� ��� ���� �����������).
� ��� �� �� ������� ����� �������� ����� ������
���, ������� � ������������ ������� ��������
����������� ������� � �������� �������� �����
���� ����������. ��� �� �����, ��������� �����
�������������� �������������� ������������
����������� ������� ������� �������.
������ ����� ����������, ��� �����������
������� � ������� �� ������� � ��������� �����
������ �� ������������������� ������� �
���-������ �� ����� � ������ (���. 12). ����������
�������� ��� ��������� � ���������� � �� ����� �
� ������-�������� � ��������� �����������,
�������� ������� � ��������. ��� ���� �����
��������, ��� ����� ������������� ����� �����
����� � ���������� ���������� �������� �������,
��� ����������� ������� � ������������ ����
�����������: ���������� � ����������
�����������, ����� �� ��� ������������ �������.
�����, ��� � ������� �������, �������
������������ �������. ������� �� �� �����������
���������� ��� ������� �� ������������ ��������
������������ � ��������� �����.
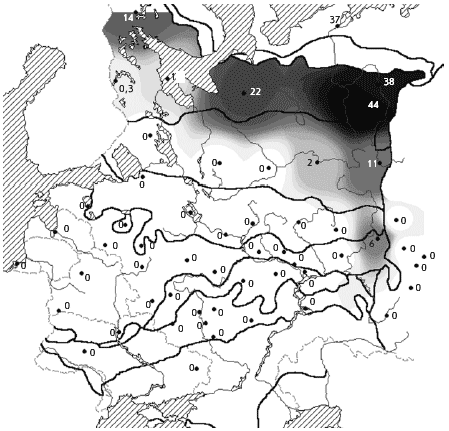
���. 12. ������. ������ ����������� �
������� � ��������� ����� (� �������), ������/��2
���� ������������� ������ � ������� ��
��������, �����������, ��� ����������� ��������
������ � ������ �� ��. ��� ������������� �
�������� � ������� �����, � ����� ����� �
���������� ����� ������, ��� ����� � �����.
��������� ����� ����������� � ������ �� ������
�������������� � ������� �������� � �������
���������� �����. �������� ���� �������� ��� �
��������, ��� � �������� ������� �������
����������� ���.
����� ��������� ������ ������������� ������,
��� � ������� �� �� ������� ������, �������� ����
� �������� ��� (����. 10). � �������� � �������
����� ��� �������� ����� ����������� � � �����
������� ����, ��� �������, ������, � ������������
������� ��� � ���� �������, � ����� � ��������
������������ ������� ����� ������� ���������
������� � ���������� ����.
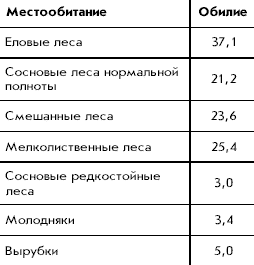
����. 10. ������������� �������������
������-������� � �������� �������� �����
(������/��2 , � ������� - ����� ����)
���
� ������ ������ �������� ������ ����� ������
������ ����� � ������, ���� ������������ ������
����������� ��������. � �������-������ ������
���� � ��������� ������� � ��������� ����� �
�������� ���.
������ ����������� �����, ��� ��, ��� � �������,
��-��������, �������� ������������ ����������
������������ ����� ���� �������� �������� ����.
��-������, �� ����������� ����� ������
���������� �� �����. ����� ���� ��������
�������� ������, ��� ����������� ����
����������� � ������ ����������� ������. �����
����, �� ������ �� ������ �������� �� �������, �
����� ���������� ������ ����� ������
������������� ��� ����, ����� ����� �������
����������� ����������� ���� �����. �������
������������ �������� ����������� ������
�������� �������������� �������, ������ �����,
�������� ����������� �����������.
������� ����������� ����� � ������� �� �����
���������� � ����� ������� � ����� ����� �������
����� ����� � �� ������� ���� (���. 13). � �������
����� ���� �����, � �������� � �� ��������.
������ ������ ������������� ����� ������������:
������������ ���������� ���������� �
����������� �������. � � ������, � � �������
����������� �����������.
���. 13. ���. ������ ����������� � �����
������� ���� (� �������), ������/��2
� ����� � ���������� ����� ��������
�������������� ����� ������ ���������; �
���������� � ��������� ����� ����������� � 3-4
���� ����, ������������ �����, ���������� �
�����-���������� �������� (����. 11). � ��������� �
����� ���� �������� ��������������� �
���������� �����, ���, ������, �������� ��������
������. �������� ����� �� � ���� ����� � �
�������� ��������������, ��� ��� ��-��
���������� ������ �������� ����� ����� ��������,
��� �� ������.
����. 11. ������������� �������������
���� � ������� (������/��2 , � ������� - �����
����)
�������
�� ����� ������ �������� ������� �����
������ �� �����. ��� ��, ��� � ����, ��� ��������
�������� �����, ������ � ��������. ��� ��
�������� ����� �������� ������ � �������
������������ � ������� �����. �� ������ �
�������� ���� �������� ����� ������� �� �����
������ ������, � ������ ����������� �������, ���
��, ��� � �����, ������������ �� ���� ���������
��������� �������� ������ � ������������
��������� ����������� �� �����. ������ �
��������� ���� � ������� ��������. ���, �����
������� ���������� �� ����� � ��������������
�������������� � ��� ������� ��� ��������������
����������� ��� �������� ����� � ����������, �
������� ������� � ��� �� ������ ����� �������
������������ ������������� ���� ����.
����� ���������������� ������������� �������
����� ���������� �� ������������� �����,
�������� �� �������� ������ �����. �������
����������� �� ���� ������������� ����������
������������������� ������� � ����� (���. 14). �
�������� ����� ��� ������; �� ������� ����� ��
���������������� ����� ���� �������������; �
��������� � ����� ������ ����� ���������. �
���������� ����� � ��������� ��������������
��������� � ����� ����������� � ���������
�����������. ���, ����������� ����������
�������� � �������� ������� (����� �������
�����������, ����� ��� ������), � �� ���� ������
����� ������� ����������� �������������.
���. 14. �������. ������ ����������� �
����� ������� ���� (� �������), ������/��2
������������� ������� �� �������� �� ���������
� ������ ����� ��������� (����. 11). � ������� ��
�������� ��� ������������ ��������� ����������
� ��������� ����, � ����� ���� � �����������. �
�����, ��� ��, ��� � ����, ������� ���� �����
�������� � ����������, ������ �������� ����� ��
�������� � � ������ ��������������. �
������������ � ��������� �����, ���������� �
�����-���������� ������� ���� �������
�������������, � ��������������� ����� � ��
������� ������ ���� ������� ������ 10 ������/��2 ;
������ � �������� �������� ������ ����������
�����������. � ����� ������ �������� �������
������������ � �����-���������� � ����������;
��������, ��� ��������� ������� �����������. ��
���� ��������� ������ ��������, ����� ��������,
������� ����� �����. � ��������� � ����� �������
���� ����� ����������� � �����, ������ �
���������� ����� ������� ����, ��������
��������.
�������
� ������ ����� ������� ���������� ����������
������� ����� ��������� ����� �������������
����� � �������������� � ����������� �� ���
������. �������� ���� �������� � ����� ��������
� �����������, � ������ ������� ������; ������
�������� � �����, �����, ������, �������� ��
����� ������ ������� ��������; ������
����������� ��������. ��� ���������� ��������
������ ������� �������� �������.
� ������ ����� ������� � ������� ���������
���������� �������. ������, � ������ � ��������
�������������� ��� ����� ����������� �� ����
������������� ���������� �������������������
������� � ����� (���. 15). ����������� � ��� ������:
� ����� ������� ���� ������� ���� ������, ����� �
�������������. �������������� ��������
����������� � �������� �������� ����������� ��
��������; � ����� �����, ���������� ����� �
��������� ������ �������� ������� �������������
� ������.
���. 15. �������. ������ ����������� �
����� ������� ���� (� �������), ������/��2
����� ������������� ����� �������������
������� ������������ ������ � �����������,
���������� ���� � �������� ����������������
����� ��������, � ����� ���� � ���� � �����������
(����. 12). � ��������� ����� ����������� � 1,5-2 ����
����, � ���������������, ������� ����� �
���������� � � 2-4 ���� ����.
����. 12. ������������� �������������
������� (������/��2 , � ������� - ����� ����)
������� ������� �����
������� ������� ����� � ������������ ���
������������, ����������� � ������ �����
������������������� ������� ���������� �������
����������� ����������� � ��������� �������� �
������ ������������ �� ����������� �����.
�������� ������ ������, ��-��������, ������
����������� ������� ������� ������ ��������
�������� ��� � �����. �� ������� ����, ���������
�����, �������� ���� �����������, ����� �������
����������� ������� �� ���������.
�������������� �������� ����������� ��������
�������� ����� ������������ ��������. ������,
����� ������� ���� ����������� ������ �
���-������� (���. 16). ������� ������� �����
��������� ����������� ����� �� ���� ������
�������� ������������������� �������. �
�������� � ������� ����� ��� ��� ������� ������,
����� � �������������. � �������� �������
�������� �������������� ������� ����������� ��
��������. � ����� �����, ���������� �
���������������� ����� � � ��������� ������
������� ������� ������ � �������� ����� �������
� 1,5-2,5 ���� ������, ��� � ���������.
���. 16. ������� ������� �����. ������
����������� � ������� � ��������� ����� (�
�������), ������/��2
� ����� ������������� ������������� �������
������� ����� ���� �������� � ����� � ��������
������� ����� (����. 9). � ����� ������ ��������
��� ������� ����������� ����� �
���������������� �����, ��� � ����� ����������
�������� ��������������� ��-��� ���� ��������.
��������������� ����� � ���������� �����
��������.
����������
�������� ���������������� ���������
�������������� ������������ ����������� ����,
�������� � ����� ������������������� ������� �
��������� � ������ �� ��. ���� ����������� �
�������� ����������� � � ��� �� ����� � �������
������ ��� ����������� ������ � ������-�������.
����������� ������ ������������� � ��������
�����, � ����� ������� ����������� ��
�����������. �������� ����������� ������
���������� �� ��� �������� ������� ����� � ����
�����. �� ���������� 14 ����� 7 ����� ���
����������� � �������� � ������� �����, 4
(�������, ��������, �������� � ������� ������)
�������� ������ �������� �����. ������������� 3
����� � ������, ������� � ������� ������� �����
� � �������� ����� ������������� ��� ������,
���� � ��� �� ����������� ����������.
����� �����, ������������� ����������� � ���,
��������� ����������� ���������� �� ����������,
���������������� � ����������� ����. ��������
����������� � ����� �������� � ��������� �
�����, �� ��������� � ����������� �
����������������� ������, ���������� ��� 3 �����
� �������, ������� � �������� ������.
�������� ������������ ����������� ���� �����
�������� �� ��������� � ���������. ����������� 7
����� (���������, ���������, ������� ������,
������������ ������, �������, �������� ������,
������) ������� ������������� � ������� �� �����.
� ���������� � �� ����� ��� ����������� ���
�����, � �������� ������� ������� �
�������������. ������������ ���� ����������� �
������� ������� ��� 3 ����� (��������,
������-������� � �������� �������� �����),
����������� ������������� ������ � ���������� �
�� �����. ��� � 4 ����� (�������, ����������,
����������� ������ � �������) ����������� �
�������� ����������� �������� �����
�����������, ����������� ����� ����������
��������� ��������� � �������. ���c��������
������ ������� � ���� �������� � �����������
�������� �������.
�������� ������� �� ����������� �����������
������������ ��������� (��������� �������������
��������) �������� ������ ��� �������� ������ �
������������ ������. �������� ������
������������ ��������� ���������� ����, �
������������� �� ������� �������� �����. ���
������������ ������ ������������ ���������
���������� ������� �� ������� ��������
����������������� ��� ������-�����������������
���� � ����������. � ����������, ��
��������������� ����������� �����������, ���
���� ����� ��� �����������. ��� ���� �� �����, �
����� ��� �������, �������������� ��������� ����
� ����������� � ������� ����. �����������
��������� �����, � ��������, ���� ������� ��
���������, � ������������ �������������
�������������� ���������� ��������.
�� ��������� �������������� ��������
������������� ���� ����� ��������� ��
����������, ������������� � ����������. �
���������� � �����, ����������� ������� ����
������� �� ������� � ������� ��������� ����� �
��� � �� ��������� �������������� ������������
��������� ������������ �������, �������� ������,
��������, �����-������ � ������� ������� �����.
��� ���� ����������� �������� �����, ��������� �
������� ������� ������ � ����� � ������ � � ����
����������� �� ����������, ����, ����
������������� ������������� ����������� ����� �
���������, �� ��� �������� ���� ��������� ������
���, ��� �������� ������ � �������. �����������
�������� � ������� � ����� � ���� ���� ����.
������ ��������������� ������������� ������
����������� �������� ������� �� �����, ���, �����
���, ��� ��� ����� ��� � �����, ��� ��� ����� ��
������� ����������� ��������� � ��������, ���
����� ������ � ������ ��������� � ��������
������������ �����������.
����-������������ ����� ����� ������ ��������
������������ ���� � �������� ����������������
�������� � ���������. � ���� ������ ���������, �
����� �������, ���� �������� ����, ��������� �
����������������� ��������� � ������� �������
����� ������ ���������� �����: ���������,
������� ������, ������������ ������, ���������. �
������ �������, ���� �� �� ���������
�������������� ������������� ����� ������� �
���������, ��������� � ���������� ����� ������
���� � ������ ��������� �������� (�����, ������,
� ������� ������� �����, ����� � ������), � �����
������ ������������� ������������ � �����,
���������������� � ����������. ��� ���� � ���,
������� � ������� � ��������������� �������
������� ����� � �����-����������, ���� ��
���������� ������ ��������� �������� � ��������.
����������- ����������� ������ �����,
����������� ������� ���������� ������ ��� �
�������, ��� � � ���������� �����. � �����
�������, ��� �����, �������������� ���� �
�������� ������� ��������, �� ���������� �����
��������� ���������� ���� �� �� �����, �
���������� ��������� ������������� � ������� �
���������� ����������. ������ ������ �
����������� ������. � ������ �������, ���� �����
������� ����, ��������� ��� � ����� � ��������
�����, ��� � � ���������� ����� � ���������� �
������.
����� ��������, ��� ����� �������� ��������
���� ��� �����, ������� ������������ ��
��������������� ���� � ���������. ��� �������
��� ���� ���� �������� ��������������� ���
�������������.
����������
Handbook of the Birds of Europe, the Middle East and North Africa .
Sen. ed. C.M. Perrins. V. 7. Oxford-New York. 1993.
������ �.�. ��������������� ��������� �
�������� �������� ������ (Parus cristatus L.) // ������.
�������. �����. �� ����. �������� ���� �� ���л. �
14. 1983. �. 43-49.
��������� �.�. ��������� � ����������
���������������� ��� �����, �������� � �������
���� � ����� ����������� // ����. ����. �. 65. ���. II.
1986. �.1664-1674.
������� �.�. ��������� ����������
���������� ���� � ���������� ����� ��� ���������
������������� ������������ ����� // ����� �
�������� ���� � ������������� ������� ������ //
�.: �����. 1987. �. 108-141.
�������� �.�., �������������� �.�., ���������
�.�. �������������� ������������
���������������� ��� ����� �������� ���� ����� //
��������. �4. 1988. �. 34-38.
����������� ������ ������ ����
����������� ����� ���л . ������ ����� 1987/88.
���. 1. ����. �.�. ���������, �.�. ��������, �.�.
������, �.�. ��������������. �. ���-�� ������. 1990.
30 �.
����������� ������ ������ ����
����������� ����� ���л . ������ ����� 1986/87.
���. 2. ����. �.�. ���������, �.�. ��������, �.�.
������, �.�. ��������������. �. ���-�� ������. 1990.
30 �.
����������� ������ ������ ����
����������� ����� ���л . ������ ����� 1987/88.
���. 3. ����. �.�. ���������, �.�. ��������, �.�.
������, �.�. ��������������. �. ���-�� ������. 1990.
32 �.
����������� ������ ������ ����
����������� ����� ������ � ������������
�������� . ������ ����� 1989/90. ���. 4. ����. �.�.
���������, �.�. ������, �.�. ��������������. �. 1994. 41
�.
����������� ������ ������ ����
����������� ����� ������ � ������������
�������� . ������ ����� 1990/91. ���. 5. ����. �.�.
��������������, �.�. ���������. �. 1995. 47 �.
����������� ������ ������ ����
����������� ����� ������ � ������������
�������� . ������ ����� 1991/92. ���. 6. ����. �.�.
��������������, �.�. ���������. �. 1995. 33 �.
����������� ������ ������ ����
����������� ����� ������ � ������������
��������. ������ ����� 1992/93. ���. 7. ����. �.�.
��������������, �.�. ���������. �. 1996. 44 �.
����������� ������ ������ ����
����������� ����� ������ � ������������
��������. ������ ����� 1992/93 � 1994/95. ���. 8-9.
����. �.�. ��������������, �.�. ���������. �. 1996. 60 �.
����������� ������ ������ ����
����������� ����� ������ � ������������
�������� . ������ ����� 1995/96. ���. 10. ����. �.�.
��������������, �.�. ��������, �.�. ���������. �.
1997. 52 �.
����������� ������ ������ ����
����������� ����� ������ � ������������
�������� . ������ ����� 1996/97. ���. 11. ����. �.�.
��������������, �.�. ������, �.�. ���������, �.�.
���������. �. 1997. 46 �.
������ �.�. � �������� ����� ���� ������
���������� // ������� ������ ���������
���������� �� �����. �����������: �����. 1967. �. 66-75.
***
������ ������ ���� ������������ � ������� �������������� ������ ������������� �����
"������� ����"
������ ����������������� ������, ��������� � ������������� ������ ����, ����� ����������
� ������� ����������������� ������ ���
������� ������
"�������� ����"
��������� "Parus"
������ ���������
���� ��������� ������� � ������������ �� ����������� � ������ ������ ��������-��������
Google Play ���
AppStore
������������ ���� ������ ,
���������� �������� � ����������� 206 ����� ���� ������� ������ (������� ����, �������, ������, ���� � ������), � ����� ������������ ��������� ����������� ����������� � ������� ����.
����� ������� ������ ,
����� �� ���� ������ �
������� ,
������ ���� ������ �
����� ������ ,
������ ���� ������ �
����� ������
(�� ����� ������� �� ��������� ���������� �� ������� �� ��������� ��������),
����� ������ ,
������������ ���� ������ (Birds of Europe) ,
������ ���� ������ ,
����� �� ���� ������
� ���� �������� ������� ,
������������ ������� ���� �
������������ ���� �� ����
(��� �� ����� ������� �� AppStore )����� ������� ������ (160 �����) ,
����� ������ (278 �����) ,
���������� ����� ,
�������� ����� ,
����� ,
�������� �������� ,
������ ���� ������� ������ ������ (343 ���� ����).

 ���� �����
���� �����

 ������������ �������� (��� PC-Windows)
������������ �������� (��� PC-Windows)