����������:
1) ������������� ����������������� ������ ��������� �������� �� ������ ������� ������������ � ��������� �� ����� � ������������ ���� (���������� �����������, ������� - �.�. ���������);
2) ����� ����� �� ����� ��������������� �� ������������� ��������, ���������� � ������;
3) ������ ������ �������� ��������� � �� ����������� �������� ������ � ��������� ������� � �������� ������������ �� ���� www.ecosystema.ru
������ ������ ���� ������������ � ������� �������������� ������ ������������� �����
"������� ����"
��������������� ������������ ����
�� ������ CAREX
SALINA WAHL. (CYPERACEAE) �� ��������� ������ ����
�.�.���������, �. �. �������
2005
��������� ����������� ����, ����������� �
������ Phacocystis (��. ���. 1) ������� � ����������
�������������, ������� ��������� �������� �
������ ��������� ����������� (�������, 1976;
�������, 1999; Duman, Kryszczuk, 1958; Haines, 2000). Carex salina, ��
������ ������ �������, �������� ����� ����������
������������� (����������������� ��������) ��
Carex subspathacea Carex paleacea (Haines, 2000; Standley, 1990; Cayouette,
Morisset, 1985). ��� �������� ������������ �� �����
���������, ��� ����������� �����, ����������
��������� �������� (allozyme), �������������
����������, �������� ���������������. ���������
������ ������������ ����� ��������� ������� Carex
recta (Carex paleacea Carex aquatilis). ���������� ���� ������,
��� Carex salina � ������� �� ����������� Carex recta Carex
subspathacea (�������, 1976).
���� ����������� ��������, �������������� �
������ Phacocystis � � ������ �������, ������ �����,
�������� �������������� ��������� �������
���������, ������� ���� ���������, ���� �����
���������� ������������, ��������� ������� ��
������ Phacocystis ���������� ������������������ �
������ �����������������. ��� ��������������
�������� �� �������� ������� ��������� � ����
�������������� ������ (Haines, 2000; Standley, 1990).
�������������� ��������� ������� �����
����������� �� ������ ������������� ����������.
�������������� �������� ����� ����� ���� ������
����� ������ ���, � ��������� �������, �����
����������� ������, ����������, �� ������
��������� ��������������, ������ ��������������
� ��������������� ����������, ��� ������� ���
������������ (Whitkus, 1988). �����������������
������� ��������� ����������� ������������
����������� � �������� ��������� �������������
����. �������� ������������ ���� ���������� �
�������� �������, � ���������, ��-�� ���������
�������������� �������� (Whitkus, 1988).
���� ������ ������ ������� � ��������� �
�������� �������� ���� ������ Carex salina, �������
��������� "� ����" �� ��������� �
������������� ��������������� ���������.
��������� � ������
� 2000, 2002 ��. �� ��������� ������ ���� (���� ����,
���� ���, ����-������� �-�� � �-�� ����������
����������) �� ��������� ���������
��������������� ��������� (��. ���. 1) ���� ����
��������� ����� (����������� Carex paleacea ����
������� ������ ����, ����� 7, � ���� �� ��� ���
��������� ���������� �� ���� ��������� �����
(�������, 1999), ������� ��� ������ ���� ���������
�� ������������).
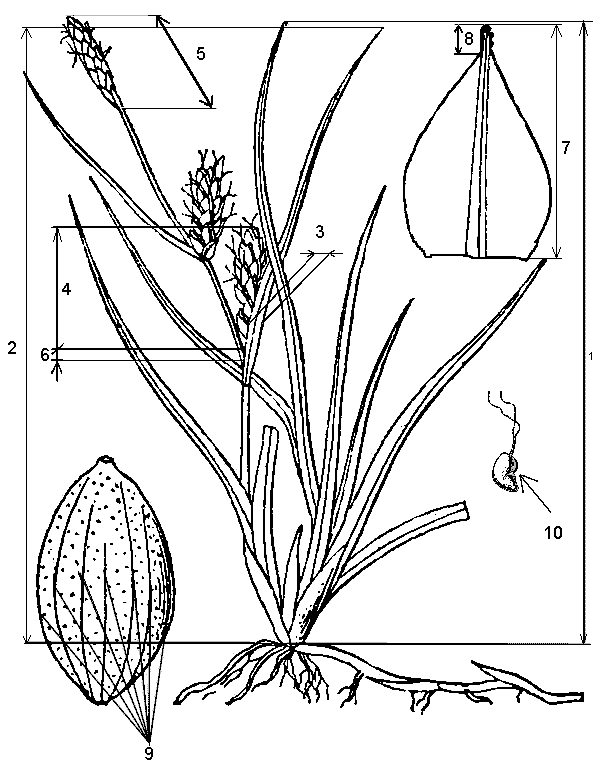
���. 1. ���������� �������� ����
����. 1. ����������� ���������:
1. ����� ������������� ������,
2. ����� ������������� ������,
3. ������ ������� ����������� ����� � ���������,
4. ����� ������� ���������� �������,
5. ����� �������� ����������� �������,
6. ����� ��� �����,
7. ����� ���������� ����� ���������� �������,
8. ����� �� ����,
9. ������� ����� �� �������,
10. ������� ���������� �� ������� �����������
�������,
11. ������������ ����������� �����. |
� 2003 �. ���������� ������� �� ��������� ����
����, �������� �� �������� ���-���� ������ ����.
��� ���������� 31 ������, ������ ����� �� 5-�
�������� ���� ��������� ����� ������ Phacocystis (Carex
aquatilis). ����� ���� �������� 671 �������� �� 28
��������� �� ��������� 11 ���������������
��������� (��. ����. 1).
��������� �� ���� � ������ ���������,
�������������� � 2000 �., ������� ���� ���������.
��� �������� ������������� ���� � ������, ������
���������, ����������� ����������� ���������, �
����� ���������, �������� ������� �� ��������
������� ��������� ���� �������������. �����
����� ��������� ���� ���������� �������, �������
� ��������� ��������, ����� �������, ������
������ ������� � ������ ������������� ����� ��
������������ ������, ���������� � ���������.
��������� ����������� �������� � ����� ������� 1
��.
��������� ����������� � ������ STATISTICA 5.5 (Statsoft,
Inc., 1999), � ����� � ����� R 1.8.1 (R Development Core Team).
���������� ������ ������� ���������, ����������,
���������������, �������������� ������ �
��������������� ������ ��������, � �����
�������������� ������������ ���������� (�����,
1984; Mathsoft, Inc., 1999).
����������� ������ �� ������������
������������� ������, ������� ��� �������
������������. �������� ����� ������������
������������ �����.
������� ��� �������� ��������������� ������. �
���� ���� ���������� ������� 2000 ���� (75 ������)
���� ������� �� ���� � ������������ � ������
(�������, 1999). ��� ���������� ����������������
������� � ���� ������ �������� �������������,
������� ����������� ���������� �� ��������,
�������� ����� ����� ������� (�� ����. 2).
����. 2. ������� ������������� ������ 2000
���� �� 3 ������ � Carex recta (I), Carex salina (II), Carex subspathacea
(III)
(������ � ����������� �������������, ������� �
�������������� �������������)
| ������ |
������� ������������ |
I |
II |
III |
| I |
87,5 |
21 |
2 |
1 |
| II |
72 |
3 |
18 |
4 |
| III |
92 |
0 |
2 |
23 |
| ��� ���� |
83,78 |
24 |
22 |
28 |
���� ����������� ������� "����������" ��
���� ����� ���������� ����� (Carex salina, Carex recta, Carex
subspathacea) ��������� ������ � Carex recta � Carex
"salinasubspathacea" ("���", � �������� ��������
�������������� ������������� Carex salina, � Carex
subspathacea) ��� Carex "salinarecta" � Carex subspathacea. �����
���� ��������� ��������� ��������� ������������
�������������, ���������� ��� ������
���������������� �������. �������� ����������
�������� ��������� � Carex recta, Carex "salinasubspathacea",
��������� � ���� ������ ����������� ����������
����� ������� ������������ � 90,54 (��. ����. 2 � 3).
����. 3. ������� ������������� ������ 2000
���� �� 2 ������ � Carex recta (I), "Carex salina-subspathacea"
(II)
| ������ |
������� ������������ |
I |
II |
| I |
83,33 |
20 |
4 |
| II |
94 |
3 |
47 |
| ��� ���� |
90,54 |
23 |
51 |
��� ������ ����� ���������� ��������� �������
(box & whiskers plot) �� ����������� ��������� (c�. ���. 2).
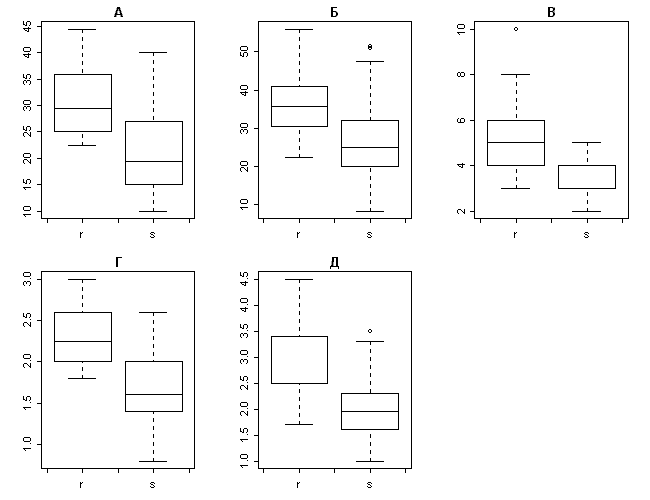
���. 2. ��������� ������� �� �����������
��������� ��� Carex recta � Carex salina s. 1.: (�) � ��.
������������� ������, (�) � ��. �������������
������, (�) � ��. ���������� ����� ����������
�������, (�) � ��. �������� �������� �������, (�) �
��. ������� �������� �������.
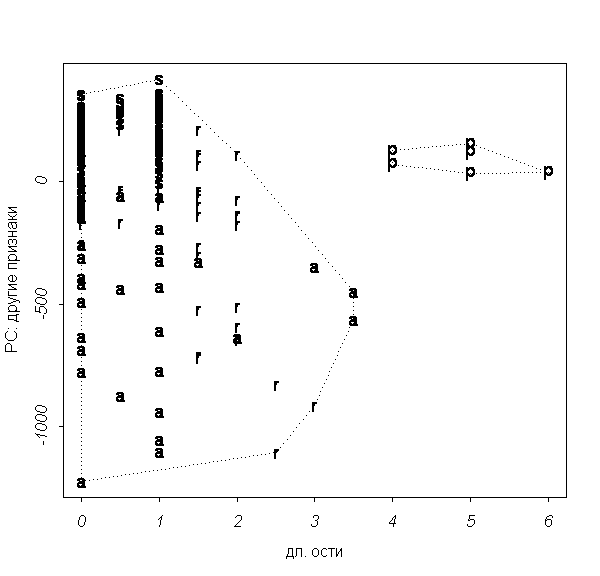
����� ��� �������� ������ ������ 2002 ����. � ����
���� ������ ���� ���������� ��� ������ �������
������� ��������� (��. ���. 5, �). � ����������� ��
���� ������� ��������� ������� �����������
������������ ����� ����. ��� ���� ����������
�������� �� ������� ���������� ����� �����
��������, ��� ����� ������������� �
������������� ������. ������ ��� �������� �����
������ ������������� �� �������� ��������� ���
������ ���������� ������� �������������������
�������������. �������, ���� ��������� ��
������������ ��� ��������, �� ���������� ������
������ (��. ���. 5, �).
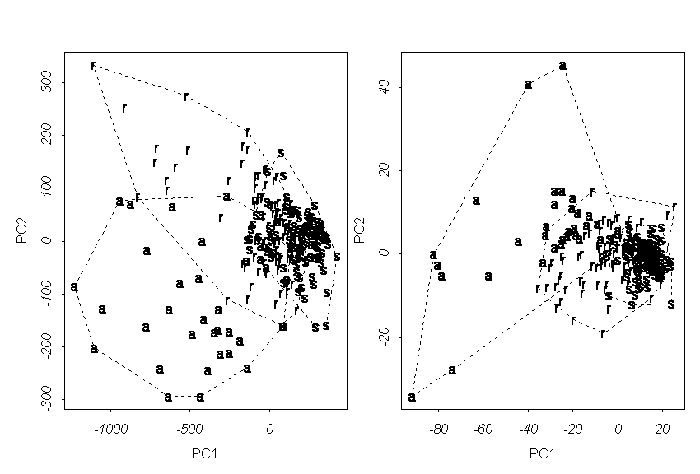
���. 5. ���������� ������ 2002 ���� �� ����
("�" � Carex aquatilis; "r" � Carex recta; "s" �
Carex salina s.l.):
(a) �� ���� ���������, (�) �� ���� ���������, �����
���� �������.
����� ��������, ��������� � Carex aquatilis �����
��������, �������� ����� (�������, 1999), ���
�������: Carex aquatilis subsp. aquatilis � Carex aquatilis subsp. stans. ��
����������� ��������� ��������� (scatterplot) �� ����
������� ��������� �������� ���������� ���� ����
�������� (��. ���. 3).
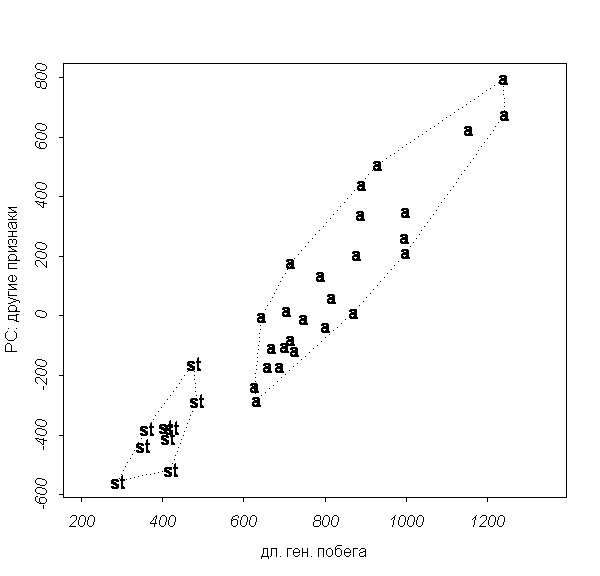
���. 3. ���������� Carex aquatilis subsp. aqatilis � Carex
aquatilis subsp. stans ("a" = Carex a. subsp. aquatilis; "st" = Carex
a. subsp. stans).
��� �������� 2003 ����, ��������� �� ���������
��������, ���� ��������� ���������� �� ���������
�� ��������� ��������� �� ���� �������
���������; ����� ������������� � �������������
������ �������������� (��. ���. 4).
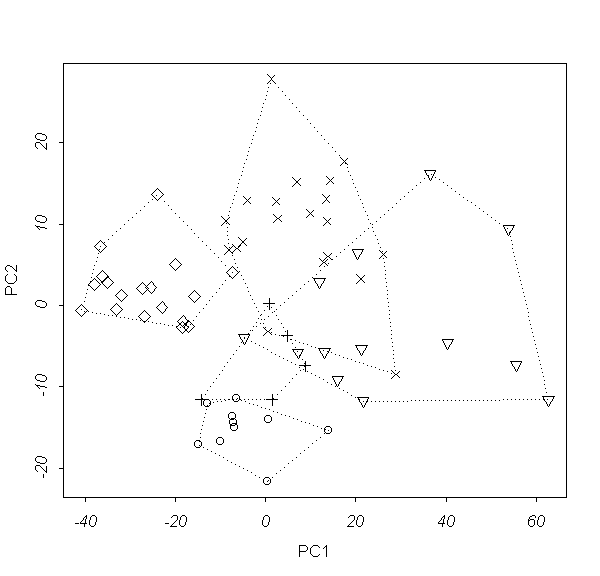
���.4. ���������� ������ 2003 ���� ��
��������� (������ ������� ������������� ������
����������).
��� ��������� � ������� ���� ����������������
������� ����������� Euphrasia, Achillea � Carex serotina(���
���� ����� �������� ����� ���������������
(Blackstock, Ashton, 2001; Pykala, Toinoven, 1994; Swann, 1983; Yeo, 1978)). ��
������� (��. ���. 7) �������� ���������� ����
�������� �� ���������� �� ���� �������
���������.
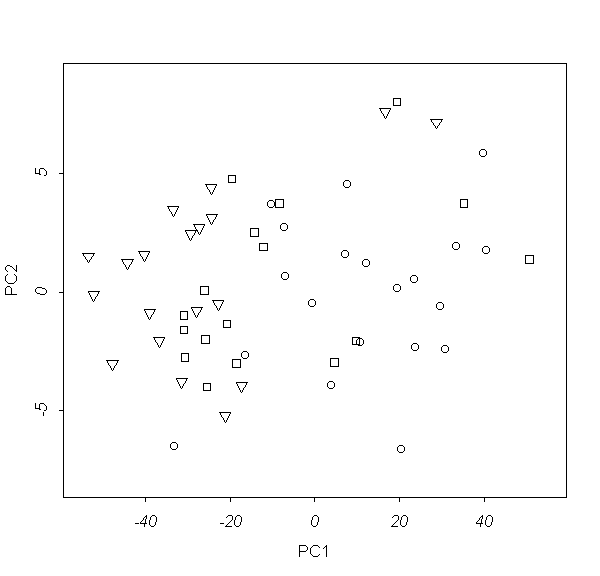
���. 7. ���������� Euphrasia �� ���������
(������ ������� ������������� ������
����������).
���������� � ����������
�� ��������� ���������� ����� ����, ���:
- Carex paleacea, �������� �� ������������, �����
���������� �� ��������� ���� ������ ��������
������;
- Carex aquatilis ������������ � ���� ���� �������� � Carex
aquatilis subsp. aquatilis � Carex aquatilis subsp. stans (��. ���. 3);
- �������� ���������� ��� ��������� ���� ��
������ Carex salina �������� ��������� ���� �����: Carex
recta � Carex salina s. l. (incl. Carex subspathacea).
��� ����������� ������� ��������������
���������, �� ������ ������������ ���������, ���
��������� ��������� ����:
1. ���������� ������������ ����� ������� 23,5 ��;
������ ������������� ������ ������ � ����������
������ ��������� 30,5 ��; ������� ������� �������
����� 1,8 �� ��.; ������ ������� ������� ����� 2,4 ��
��.; ������� ����� 5-6 �� ��., �������� Carex recta
������� ������� � �������� ������ �����������
� ����, ������� ����� 3-4 �� ��., �������� ��� ���
���� Carex salina s.l. (incl. Carex subspathacea)
���������� ��������� ��������, ��� ������
��������� ���������� ���� �� ����� ����������,
��� ����, ������ ����������� ����� ���������
�����������, ���� ������ �� ������������ �����
��������, ��� ����� ������� (��. ���. 4 � ���. 5).
��������� ��������� ������ ���������� ��������,
���������������� ��������������� ���� ������
Phacocystis ����� ��������������� ��� ������
������������� � �������� ��������. �����
�������� ����� ����� ����� ��������� ����,
�������������� �� ������ ��������.
� ������� �� ��������� Achillea, � ������� ��
��������� Euphrasia ������ ����������� � ������
��������� ����� �����������. ��� ��������
������������ �� ������� ��� ���������, �������
������������ ��� ��, ��� ���������������
��������� ���� � ����� ����������� ����� (��.
���. 4 � 7). ������ �����, ��� ������� � ���, ���
������� ����� ������� (��������� ���������)
����������� ��������� � ���� �����
���������������� ��������. ���� ���������
��������� ������ �� ����� ���� ���������;
�������� ��� ����������� ��������� ���������
�������, �� �� ���������, ��� ����������������
�������� ������������� ���� � ����, ��� � ������
���������� �����.
������
������ ����������� ��� ����������
�������������� ���������������� �����������.
�������������
�������� ������������� ��������������� ��� �
������ ��������� �., �������� �., ������� �.,
�������� �., �������� �, ���� �., �������� �.,
��������� �., �������� �., ��������� �.,
�������-���������� �.
����������
- ������� �. �. ����� (Carex L.) ������ � ������������
����������. ���., 1999. Carex 426-430.
- ������� �. �. ����� ����������� ����� ����. ���.,
1976. Carex 197.
- ����� �. �. �������������� ������ � ��������.
���., 1984. 288 �.
- Blackstock N., Ashton P. A. A re-assesment of the putitave Carex flava agg. (Cyperaceae)
hybrids at Malham Tarn (v.c. 64): A morphometric analysis // Watsonia. 2001. Vol. 23. P.
505-516.
- Cayouette J., Morisset P. Chromosome studies on natural hybrids between maritime species
of Carex (sections Phacocystis and Cryptocarpae) in northeastern North America, and their
taxonomic implications // Can. Journ. Bot. 1985. Vol. 63. P. 1957-1982.
- Duman M. G., Kryszczuk S. D. Introgressive hybridization in the Carex stans-bigelowii
complex // Bull. Torr. Bot. Club. 1958. Vol. 85. P. 359-362.
- Haines A. Identification and taxonomy of two difficult maritime hybrids with Carex
paleacea // Botanical Notes. 2000. N. 4. [Electronic resource]. Mode of access:
http://www.woodlotalt.com/publications/ publications.htm.
- Pykala J., Toinoven H. Taxonomy of the Carex flava complex (Cyperaceae) in Finland //
Nord. J. Bot. 1994. Vol. 14. P. 173-191.
- R Development Core Team. R: A language and environment for statistical computing // R
Foundation for Statistical Computing, Vienna, Austria [Electronic resource]. Mode of
access: http://www.R-project.org.
- Standley L. Allozyme evidence for the hybrid origin of the maritime species Carex salina
and Carex recta (Cyperaceae) in Eastern North America // Syst. Bot. 1990. Vol. 15. P.
182-191.
- Saukel J., Langer R. Die Achillea millefolium-Gruppe (Asteraceae) in Mitteleuropa //
Phyton. 1992. Vol. 31. P. 185-207.
- Schmid B. Notes on the nomenclature and taxonomy of the Carex flava group in Europe //
Watsonia. 1983 Vol. 14. P. 309-319.
- S-PLUS 2000 Guide to Statistics // Data Analysis Products Division MathSoft, Inc. 1999.
Vol. 2. Seattle, Washington.
- STATISTICA for Windows [Computer program manual] // StatSoft, Inc. 1999. Tulsa, OK.
- StatSoft, Inc. (2001). ����������� ������� �� ����������.
������, StatSoft. WEB: http://www.statsoft.ru/home/textbook/ default.htm.
- Swann E. L. Norfolk Euphrasiae // Watsonia. 1983. Vol. 9. P. 345-356.
- Whitkus R. Experimental hybridizations among chromosome races of Carex pachystachya and
related species Carex macloviana and Carex preslii (Cyperaceae) // Syst. Bot. 1988. Vol.
13. P. 146-153.
- Yeo P. F. A taxonomic revision of Euphrasia in Europe // Bot. Journ. Linn. Soc. 1978.
Vol. 77. P. 223-334.
***
������ ������ ���� ������������ � ������� �������������� ������ ������������� �����
"������� ����"
������ ����������������� ������, ��������� � ������������� ������ ����, ����� ����������
� ������� ����������������� ������. ������ ������� ����������������� ������ (����������������� �������), ����������� �� ����� �����, ����� ����������
���.
 ������� ������
"�������� ����"
��������� "Parus"
������ ���������
������� ������
"�������� ����"
��������� "Parus"
������ ���������
|


 ������������ �������� (��� PC-Windows) ������������
������������ �������� (��� PC-Windows) ������������
